



18/09/2016
Comment le spermatozoïde déclenche la fécondation de l'ovule
Comment le spermatozoïde déclenche la fécondation de l'ovule
par Damien Mascret, Figaro.fr Santé du 29/08/2016
Drivée par Christine Gourier, une équipe de physiciens du laboratoire de Physique Statistique (LPS) de l'École normale supérieure de Paris (ENS-CNRS) a pu récemment mettre en évidence que c'est un mode particulier de battements du flagelle du spermatozoïde qui déclenche la fécondation.
C'est l'instant crucial où le spermatozoïde atteint enfin sa cible puis fusionne avec elle. Le début d'une nouvelle vie. Une délicate prise de contact entre le gamète mâle et le gamète femelle, avant que la fusion des deux ne se produise. Deux étapes que les spécialistes de la reproduction résument en une appellation, l'« interaction gamétique ». Un moment complexe, car il ne suffit pas de mettre en contact un spermatozoïde fécondant avec un ovocyte fécondable pour que la fusion se produise.
« Il se passe deux à trois minutes entre le contact et la fusion, mais ce moment a été très peu étudié jusqu'ici, principalement en raison de difficultés techniques », explique Christine Gourier. Et de façon inattendue, ce sont les mouvements de la queue du spermatozoïde, le flagelle, qui jouent un rôle déterminant.
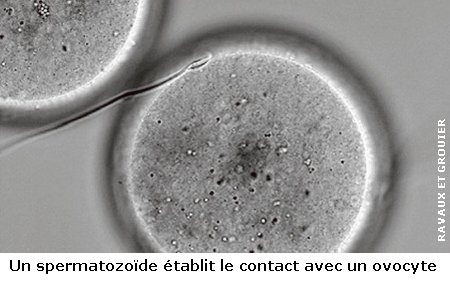
Un ingénieux dispositif pour observer in vivo l'instant crucial de la fécondation
L'équipe de l'ENS a tout d'abord imaginé un ingénieux dispositif pour que la fécondation se produise sous l'œil du microscope optique confocal. « En théorie, chez l'humain le contact peut se faire à n'importe quel endroit de l'ovocyte (et sur 80 % de la surface chez la souris, utilisée dans ce travail). Pour pouvoir observer les deux ou trois minutes qui précèdent la fusion, il fallait être sûr du site de l'ovocyte où le spermatozoïde allait établir le contact », explique Christine Gourier. Les chercheurs ont donc placé l'ovocyte dans un coquetier microscopique qui n'en exposait que 2 % de la surface, sur lesquels aboutissait un canal micro-fluidique (sorte de paille de 30 micromètres de diamètre). Dans ce canal, était introduit un spermatozoïde fécondant.
Un certain type de battement déclenche la fécondation
Benjamin Ravaux, le doctorant de l'équipe, s'est chargé des centaines de manipulations qui ont permis les découvertes. Et le spermatozoïde a fait son œuvre. « Contrairement à ce que l'on pensait du battement du flagelle, celui-ci ne sert pas seulement à conduire le spermatozoïde jusqu'à l'ovocyte, mais il sert aussi à déclencher la fécondation, détaille Christine Gourier. Et encore, seulement s'il bat selon un mode bien précis. » Les biologistes avaient déjà observé que le flagelle du spermatozoïde cessait ses battements dans les deux minutes qui suivaient le contact avec l'ovocyte, mais personne n'avait imaginé que ces deux minutes étaient cruciales. « Si on empêche ce battement, cela bloque la fécondation », explique Christine Gourier.
Pour la première fois, les chercheurs ont pu observer finement des centaines d'interactions gamétiques et identifié trois types de battements, dont un seul s'est avéré efficace pour que la fécondation s'enclenche. « C'est un battement oscillatoire assez rapide (environ 2 battements par seconde) du flagelle qui pendant deux minutes va faire osciller la tête du spermatozoïde en la plaquant sur l'ovocyte pour induire la fusion », explique Éric Perez, un des membres de l'équipe. Les mouvements amples en coup de fouet (1 par seconde) ou ceux plus réduits et très rapides (3/sec) ne permettent pas la fécondation.
Le dispositif mis au point par les chercheurs de l'ENS a ensuite permis de visualiser précisément la plongée de la tête du spermatozoïde, porteur de l'ADN issu du père, dans l'ovocyte. « Lorsqu'elle arrive à une profondeur suffisante, la masse de l'ADN paternel enfle, indiquant que celui-ci se décondense (se débobine), 50 minutes après l'entrée en contact, explique Éric Perez. Ces travaux éclairent d'un jour nouveau les trois premières minutes de la fécondation et ouvrent la porte à de nouvelles méthodes d'assistance à la procréation. »
Article original :
Benjamin Ravaux, Nabil Garroum, Eric Perez, Hervé Willaime, Christine Gourier, (2016). — A specific flagellum beating mode for inducing fusion in mammalian fertilization and kinetics of sperm internalization Scientific Reports, Nature Publishing Group, 2016, 6, pp.31886.
07:01 Publié dans Actualité des Sciences, Biologie, Sexualité-embryologie | Tags : fécondation in vitro, flagelle du spermatozoïde, déclenchement de la fécondation | Lien permanent | Commentaires (0) |  Facebook | | |
Facebook | | |
04/06/2016
Le Sénat vote contre les coraux et pour le développement économique
Le Sénat vote contre les coraux et pour le développement économique
Écoutez également l'émission de France Inter.
La France, deuxième propriétaire du plus grand patrimoine corallien au monde, a vu lundi 23 mai 2016 le Sénat retoqué l’interdiction de la destruction des coraux dans le cadre de la loi sur la biodiversité. Pourtant, grâce à ses 10 % de récifs coralliens au monde, la deuxième plus grande barrière de corail et le plus grand lagon mondial, la France pourrait être un acteur majeur dans la conservation des récifs coralliens. Conscient de cette richesse, en 1999, le pays affichait pourtant un désir de conserver ce patrimoine avec l’Initiative Française pour les Récifs Coralliens, un objectif désormais bafoué par les sénateurs.
Se disant anxieux de « ne pas contraindre le développement des économies locales », le Sénat a posé un amendement contre le projet de loi qui vise à la protection des récifs coralliens, ainsi les sénateurs consentent au dragage et à la destruction des coraux. Un tel amendement encouragera sûrement des projets comme en Guadeloupe où, pour l’agrandissement du port de Pointe-à-Pitre, 7 millions de m3 de coraux ont été réduits à néant. Un autre projet portuaire à Fort-de-France, en Martinique, a lui été entravé par les écologistes qui, après 3 ans de batailles juridiques, ont réussi à ce que les constructeurs se fournissent dans les carrières plutôt que dans les coraux pour faire des remblais. Or, avec cet amendement, les sénateurs endiguent les actions de défense pour la nature, défendant des intérêts économiques à court terme contre un écosystème indispensable qui génère des revenus d’environ 375 milliards dans le monde, chaque année.
Cependant, les coraux engendrent bien plus que des revenus économiques liés au tourisme ; couvrant moins de 0,2 % des océans, ils abritent 30% de la biodiversité marine. Bien plus générateurs de richesse étant vivant que comme remblais, ces récifs sont des nurseries et des garde-manger pour les poissons qui alimentent la pêche. De plus, si les scientifiques notent une bonne reprise du corail dans le milieu naturel, malgré qu’il s’agisse d’un processus très long, il reste essentiel de ne pas détruire les récifs déjà en danger par l’augmentation de la température de l’eau.
Et pourtant, les coraux qui subissent un blanchissement extrême n'ont ps besoin de cette prise de position pour être menacés de disparition (voir l'article de Sciences et Avenir).
13:24 Publié dans Biologie, Cnidaires, Environnement-Écologie, Limnologie-hydrobiologie, Pollution | Tags : sénat, coraux, dragage des coraux | Lien permanent | Commentaires (0) | Facebook | | |
15/02/2016
La fabrique de micro-organes humains
La fabrique de micro-organes humains
Une équipe viennoise de l’Institut de biologie moléculaire de l’Académie des sciences d’Autriche a réussi à cultiver in vitro, à partir de cellules souches humaines, un mini-cerveau de 4 mm dont l'architecture intérieure reproduit, en effet, certaines caractéristiques propres au cerveau humain.
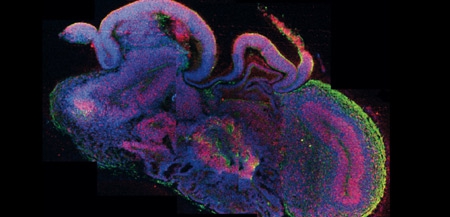
Coupe transversale d'un mini-cerveau
obtenu par culture de cellules souches
Ainsi, ces cellules sont parvenues, au bout de 20 à 30 jours, à reproduire des tissus ressemblant à un organe humain en récapitulant les mêmes processus que lors de la formation d’un embryon : elles se sont spontanément associées avant de se différencier puis de s’agencer au sein du tissu en formation.
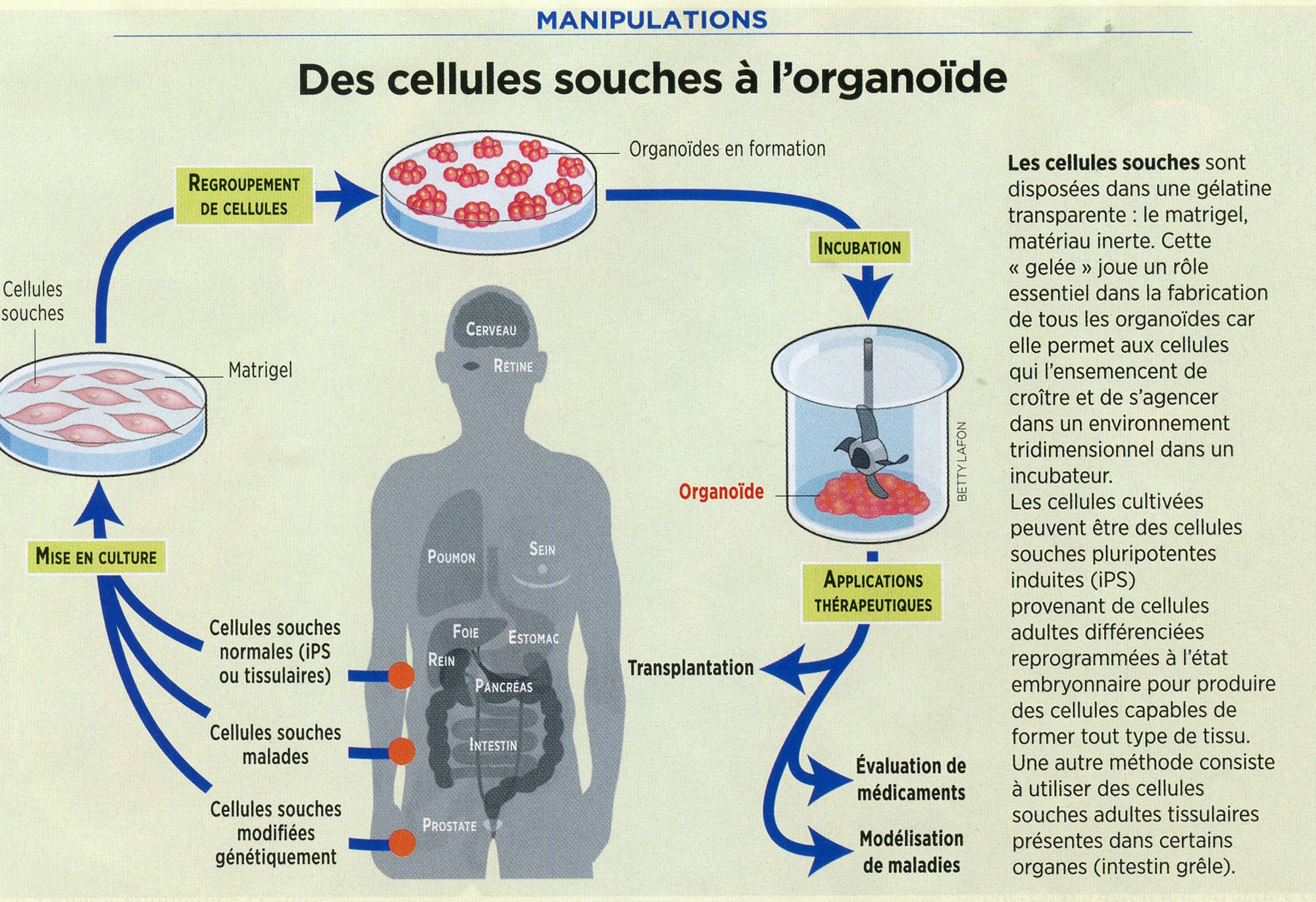
Des cellules souches à l'organoïde
Cliquez sur l'infographie pour l'agrandir (©Betty Lafon / Sciences et Avenir)
Objectif de ces recherches : tester des médicaments, servir de modèles de maladies, notamment des cancers, ou être utilisés comme thérapie cellulaire.
Source : Gozlan M. (2016) La fabrique de micro-organes humains Sciences et Avenir n° 828 — février 2016, pp. 70-72.
15:12 Publié dans Actualité des Sciences, Biologie, Médecine, Sexualité-embryologie | Tags : cellules souches, cellule souche adulte, ips, culture organotypique | Lien permanent | Commentaires (0) | Facebook | | |
09/01/2016
Cellules souches pluripotentes : une recette de fabrication qui fait polémique
 Cellules souches pluripotentes :
Cellules souches pluripotentes :
une recette de fabrication qui fait polémique
"Une annonce à la limite du malhonnête". C'est en ces termes que Mathilde Girard, chargée de recherche en modélisation pathologique iPS à l'institut I-Stem à Evry, qualifie le communiqué de la prestigieuse École polytechnique fédérale de Lausanne (EPFL). L'EPFL faisait savoir qu'une équipe de recherche était parvenue à produire des cellules souches pluripotentes induites (capables de se différencier en types de cellules qui composent un organisme adulte, exactement comme une cellule souche embryonnaire) en comprimant certaines cellules. Des travaux intéressants publiés dans la revue Nature materials mais dont l'annonce -"survendue", dixit Mathilde Girard - a soulevé un vent de critiques de la part de spécialistes francophones et anglophones des cellules souches.
Pour en savoir plus : Article de Lise Loumé sur le site de Sciences et Avenir.
17:42 Publié dans Biologie, Médecine | Lien permanent | Commentaires (0) | Facebook | | |
03/03/2015
Apparition des cellules eucaryotes (cellules à noyau) : une nouvelle hypothèse
Apparition des cellules eucaryotes (cellules à noyau) : une nouvelle hypothèse
Unité fondamentale de la vie, la cellule est au cœur de toute la biologie.
Chaque organisme complexe (plantes, animaux, champignons) est constitué de cellules eucaryotes, les cellules avec un noyau et d'autres machines internes complexes utilisées pour remplir les fonctions d'un organisme a besoin pour rester en vie et en bonne santé. Par exemple l'organisme humain comprend 220 types différents de cellules eucaryotes qui, travaillent en groupes, contrôlent tout, de la pensée et de la locomotion à la reproduction et à la défense immunitaire.
Chacune de nos cellules est composée d'un noyau et d'un entrelacs de membranes dont les biologistes cherchent l'origine depuis près d'un siècle. Jusqu'ici l'idée prévalait qu'une cellule avait grossi puis créé son noyau. Mais des zones d'ombre subsistaient.
La théorie de l'endocytose, due à la scientifique américaine Lynn Margulis (1966), expliquait déjà l'origine des chloroplastes et des mitochondries comme des bactéries endosymbiotiques capturées par les cellules eucaryotes pour se fournir en de tels organites.
En octobre 2014, David Baum, spécialiste de l'évolution à l'université du Wisconsin (États-Unis) a proposé une hypothèse audacieuse, mais très plausible. Selon le scénario de David Baum, une cellule primordiale constituerait dès le début du processus le noyau, et c'est lui qui commanderait le développement d'un nouveau corps autour de lui. Autrement dit : au commencement serait le noyau, puis viendrait la cellule.
Si l'on reprend la chronologie de l'apparition de la vie sur la Terre, on note les étapes suivantes :
- 4,56 milliards d'années : formation de la Terre.
- 3,8 milliards d'années : apparition des premières cellules simples sans noyau, les procaryotes.
- Entre 2,7 et 1,8 milliards d'années : apparition des premières cellules avec noyau, les eucaryotes.
- 1,6 milliard d'années : premiers organismes pluricellulaires (algues).
- 635 millions d'années : premiers animaux.
Ainsi, l'origine de la cellule eucaryote est considérée comme l'un des événements évolutifs les plus critiques de l'histoire de la vie sur Terre. Sans cette apparition des cellules eucaryotes, notre planète serait un endroit très différent, peuplé entièrement par des procaryotes, des organismes unicellulaires comme les bactéries et les archées.
A priori, cette chronologie ne laisserait aucun doute. Au commencement étaient des cellules sans noyau (procaryotes). Puis, parmi ces cellules primordiales qui peuplaient notre planète à l'aube de la vie, il y a 2 milliards voire 3 milliards d'années, certaines audacieuses ont tenté... autre chose. De simples, celles-ci se sont engagées sur un nouveau chemin évolutif.
Après la sobre perfection des premières, elles ont ouvert l'ère de la subtile complexité des cellules à noyau (eucaryotes), dont nous descendons aujourd'hui en droite ligne.
Non qu'il y ait eu remplacement : les procaryotes sont encore là — ce sont les bactéries, les archées, qui pullulent toujours. Mais les eucaryotes allaient inventer la pluricellularité, le sexe.
L'apparition de ces cellules d'un nouveau type a été la plus grande révolution qu'ait connue la vie... Or, on ne sait rien de cette apparition. Ni quand ni comment ni avec qui elle s'est faite.
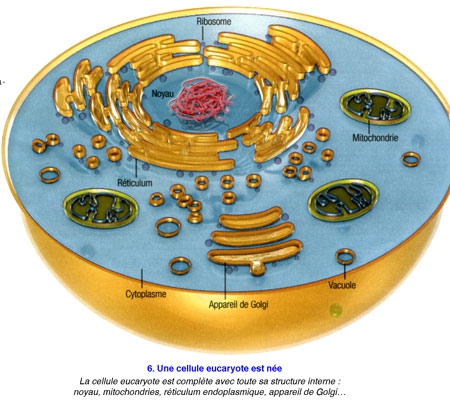
Ce qu'on sait, c'est que les procaryotes se sont munis d'une enveloppe cellulaire limitant un cytoplasme où se mêlent ADN, protéines et machineries cellulaires destinées à perpétuer la vie cellulaire. Alors que la cellule eucaryote est 10 à 100 fois plus grande et dotée d'une structure dûment compartimentée, avec un noyau central où s'insère l'ADN, tandis qu'autour se déploient un vaste entrelacs de membranes lié à la production de protéines, des mitochondries pour générer l'énergie, l'appareil de Golgi pour réguler le transport interne, un centrosome pour permettre la division cellulaire, etc.
Procaryotes, eucaryotes... Depuis qu'ils ont découvert cette dichotomie fondamentale, il y a près d'un siècle, la question taraude les scientifiques... Comment est-on passé de l'un à l'autre ? Comment est apparu le noyau dans les cellules ?
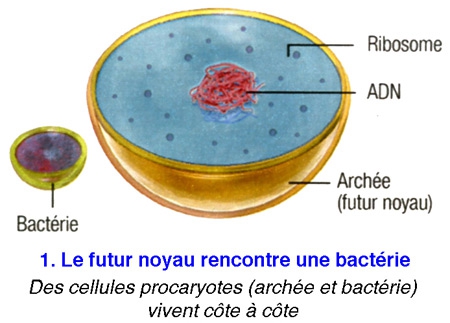
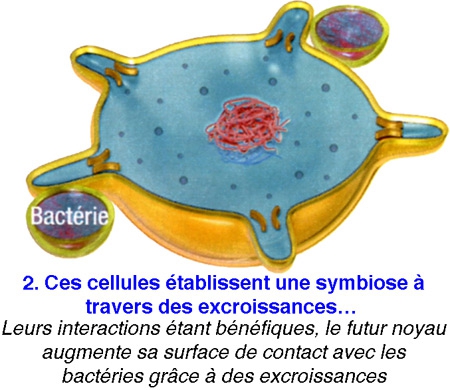
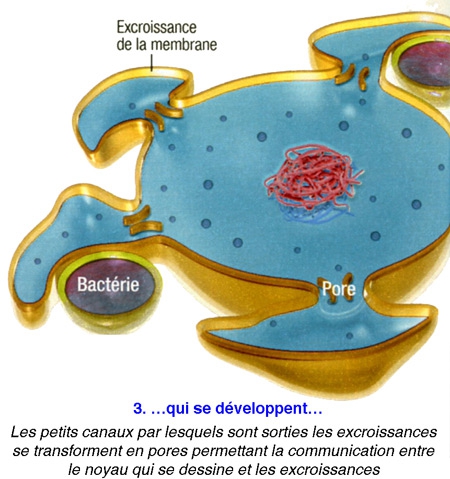
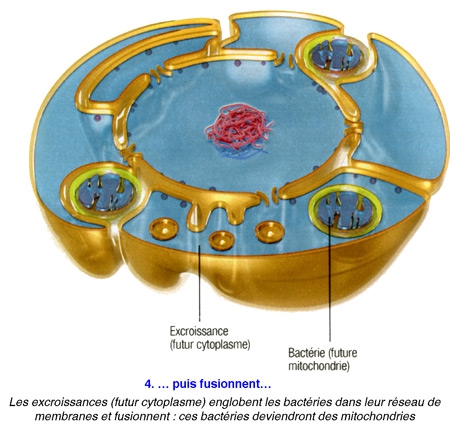
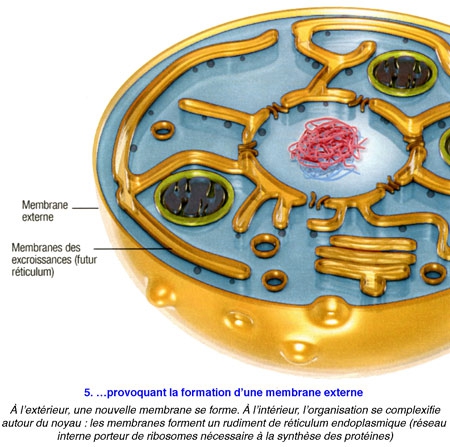
Poser ainsi la question, c'était déjà suggérer la réponse... De fait, tous les scénarios élaborés jusqu'ici sont basés sur la même trame : en l'occurrence : une cellule, sans doute une archée[1], aurait grossi en dilatant sa membrane externe,puis, à la fin de cette transformation, aurait projeté des replis internes afin de former en son centre le noyau et les complexes jeux de membranes qui l'entourent - les mitochondries étant d'anciennes bactéries avalées par leur hôte. Cela semblait logique.
La plupart des scientifiques conviennent donc que les cellules eucaryotes ont surgi à partir d'une relation symbiotique entre les bactéries et les archées.
Problème : on ne connaît pour l'heure aucun procaryote capable d'un tel trafic de membrane... Et il est difficile d'expliquer comment s'est mise en place l'organisation intracellulaire labyrinthique que l'on observe aujourd'hui.
Or, voici qu'un biologiste vient renverser toute l'histoire. Spécialiste de l'évolution à l'université du Wisconsin (États-Unis), David Baum affirme aujourd'hui que la cellule initiale — celle qui fit le grand saut pour devenir eucaryote — aurait, dès le début du processus, constitué le noyau, et c'est ce dernier qui aurait "piloté" le développement d'un nouveau corps autour de lui.
Ce scénario élaboré par David Baum en collaboration avec son cousin Buzz Baum, biologiste cellulaire à l'University Collège of London présenté pour la première fois fin 2014 a l'avantage d'expliquer, via un unique processus, l'origine si mystérieusedes enchevêtrements de membranes internes : ces derniers seraient les témoins de la naissance des eucaryotes. Connu comme la théorie "inside-out" de l'évolution de la cellule eucaryote, ce point de vue de de la complexification de la vie a été publié le 28 octobre 2014 dans la revue en libre accès BMC Biology. (voir infographie).
Infographie Science & Vie n° 1171 avril 2014
Si, dans la communauté scientifique, tout le monde n'est pas forcément d'accord avec ce modèle, on s'entend pour souligner son audacieuse simplicité et l'intérêt des réflexions qu'elle suscite sur la dynamique des cellules, le vieillissement, l'apparition des cancers...
David Baum conduit actuellement une série d'analyses génétiques pour tester justement ses idées sur ce point. Quant à son cousin, il s'est lancé dans des expériences chez des eucaryotes et des archées. Tous deux ont une série de prédictions qu'ils voudraient tester pour que le mystère de la naissance de nos cellules soit enfin résolu.
Source :
Émilie Rauscher (2015).- Cellules eucaryotes : l'hypothèse que personne n'attendait. (Science & Vie n° 1171 avril 2015, pp.74-77).
[1] Les archées et les bactéries représentent deux des trois grands domaines de la vie. Le troisième étant constitué par les eucaryotes, des organismes composés des cellules eucaryotes plus complexes.
11:05 Publié dans Actualité des Sciences, Biologie | Lien permanent | Commentaires (0) | Facebook | | |
02/03/2015
Arbre phylogénétique des Insectes
 Arbre phylogénétique des Insectes
Arbre phylogénétique des Insectes
Les insectes : 600 familles, 29 ordres, plus d'un million d'espèces ! Il aura fallu une centaine de biologistes et de généticiens pour doter enfin les insectes d'un arbre généalogique précis… qui remonte à la conquête de la terre ferme, il y a 479 millions d'années.
Ainsi, les insectes constituent le groupe animal le plus abondant sur la planète - loin devant les poissons (30 000 espèces), les oiseaux (10 000 espèces) et les mammifères (5400 espèces). Ce groupe très diversifié est caractérisé par 6 pattes (ce sont des hexapodes) et un corps en trois parties : tête, thorax, abdomen. Les araignées et les mille-pattes n'en font donc pas partie.
C'est une immense étude pluridisciplinaire d'une ampleur peu commune qui unit une centaine de biologistes moléculaires, bio-informaticiens, statisticiens, généticiens et paléontologues un travail considérable qui a permis la réalisation de cet arbre. 37 fossiles, complets uniquement, ont été étudiés. L'ADN de 103 espèces appartenant à tous les groupes connus a été analysé, et 1478 gènes codants pour une protéine selon les espèces ont été traqués dans les génomes de 12 espèces de référence représentant les familles les plus importantes : l'accumulation des différences d'une lignée à l'autre permettant d'évaluer leur lien de parenté.

Phylogenèse du monde vivant (Sciences & Vie)
Depuis le XVIIIe siècle et le premier classement du naturaliste Carl von Linné, les chercheurs essaient de comprendre leur organisation. Par esprit de synthèse, mais aussi parce que cela éclairerait l'évolution de tout le vivant tant blattes ou abeilles ont façonné les écosystèmes.
Dessiner cet arbre généalogique est un casse-tête : bien des familles ont vu leurs liens changer au fil des découvertes. D'où l'importance du nouvel arbre qui vient d'être publié. Réalisé par une centaine de chercheurs coordonnée par Bernhard Misof, du Centre de recherche sur la biodiversité moléculaire du Muséum Alexandre-Kœnig de Bonn (Allemagne), ce travail pluridisciplinaire est d'une ampleur inégalée. Surtout, comme le souligne André Nel, paléo-entomologiste au Muséum national d'histoire naturelle de Paris, “il donne un nouveau point zéro sur la phylogénie et la reconstruction de l'histoire évolutive des insectes”. Et leur interminable généalogie est enfin dotée d'une ossature solide, avec des datations et des liens de parenté précis.
En utilisant un nombre considérable d'informations contenues dans la séquence génique d'insectes, l'équipe a construit un nouvel arbre phylogénétique montrant comment ces invertébrés ont évolué et les liens qui les unissent les uns aux autres. Cet arbre suggère que les insectes ont évolué il y a environ 479.000.000 d'années, à l'époque où les plantes ont colonisé la terre. Il montre aussi que les insectes sont très étroitement liés aux crustacés cavernicoles.
Les fossiles fournissent un ancrage et une datation physiques. L'arbre phylogénétique ainsi constitué est le fruit de ces deux types dedonnées.Les insectes représentent 80 % des animaux connus ; plus d'un million d'espèces recensées, organisées en 600 familles et 29 ordres. Ils ont colonisé tous les climats, tous les milieux, et ont été les premiers à le faire. C'est le seul groupe qui a traversé l'histoire de la vie sur la terre ferme depuis ses débuts il y a 500 millions d'années.
Les auteurs de l'étude, qui font partie d'un consortium international de travail sur le projet 1K Insectes Transcriptome Evolution (1KITE), ont commencé leur travail en 2011 avec l'aide d'une nouvelle technologie de séquençage afin de clarifier les relations entre insectes étudiées précédemment en utilisant des preuves morphologiques ou des ensembles de données moléculaires plus petites. Ils ont séquencé les transcriptomes de 103 espèces d'insectes distribuées dans tous les ordres d'insectes vivants. Ils ont également exploité des données précédemment publiées des séquences du génome entier de 14 espèces d'arthropodes, ainsi que du transcriptome de 27 espèces supplémentaires. Ils ont ensuite réduit leurs données génétiques pour 1478 gènes codant pour des protéines qui sont présentes dans toutes les espèces analysées.
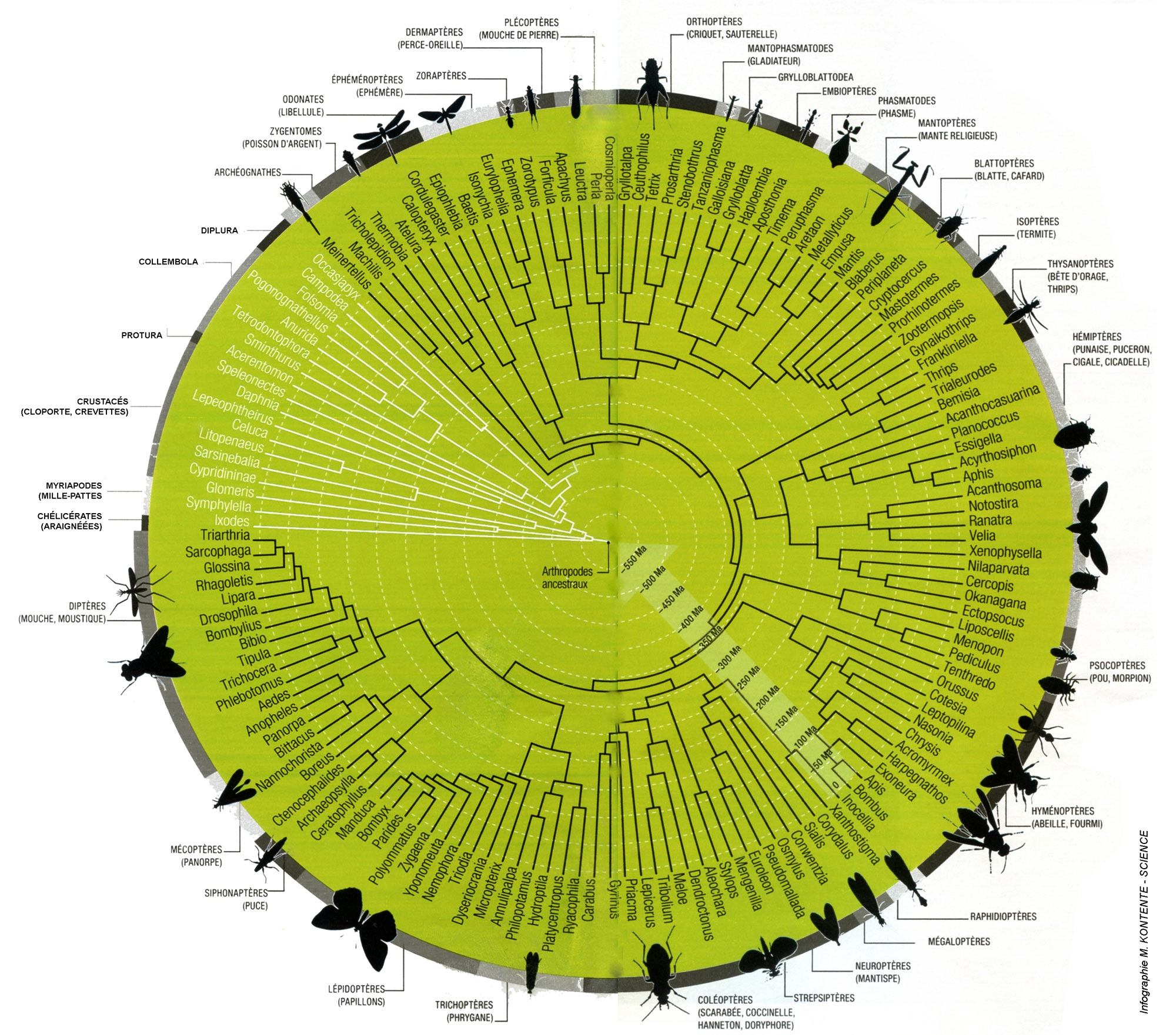
En comparant les différences et les similitudes entre les séquences de ces gènes codant pour des protéines, ainsi que les séquences d'acides aminés des gènes codants, les chercheurs ont réussi à créer un arbre montrant les relations entre 144 genres d'insectes.
Les chercheurs ont également cherché à ancrer le calendrier de l'évolution des insectes en calibrant des points sur leur arbre à partir de l'âge de 37 espèces fossiles. Alors qu'on considérait que l'apparition des insectes fossiles remontait à 412.000.000 années, l'équipe a conclu que les insectes ont quitté le monde marin et colonisé l'environnement côtier il y a environ 479.000.000 années (donner ou prendre 30 millions d'années). Cela signifie que les insectes ont colonisé le milieu terrestre en même teps que les plantes. autour lorsque les plantes ont fait. “Pour moi, la concordance entre la colonisation des insectes des écosystèmes terrestres et les premières plantes est vraiment l'une des plus importantes découvertes”, commente Jakub Prokop, un entomologiste de l'Université Charles à Prague en République tchèque qui n'était pas impliqué dans l'étude.
L'article suggère également que les insectes ont quitté le milieu marin, il y a environ 406 millions d'années, soit plus de 80 millions d'années avant que les insectes ailés deviennent abondants dans les archives fossiles, et avant que les poux parasitaires se diversifient il y a 53 millions d'années, juste au moment de l'extinction des dinosaures. Il a déjà été soupçonné que ces animaux sont apparus en même temps que les dinosaures théropodes à plumes il y a 130 millions d'années, puis plus tard en marche pour les oiseaux et les mammifères.
Pour zoomer, cliquer sur le schéma
Selon Jakub Prokop, “Les résultats de ce travail sont immenses et seront largement adoptés dans les livres et les manuels entomologie générale et systématique”.
David Grimaldi, conservateur des insectes au Musée américain d'histoire naturelle à New York, souligne que la tendance générale des relations d'insectes dans l'arbre, à quelques exceptions près, renforce la conception de la façon dont les insectes sont liés les uns aux autres. “Il est assez étonnant de voir que, après combien de gigabases de données qu'ils ont que nous ne avons pas vraiment changé fondamentalement nos points de vue sur les relations entre insectes”.
Les résultats ont confirmé de nombreuses relations, avec toutefois quelques conclusions inattendues, D'après Grimaldi, ce qui est surprenant, mais plausible que l'ordre des Diplura (insectes primitifs aptères et aveugles) n'est pas regroupé avec un autre groupe d'invertébrés primitifs semblables, les Collemboles, mais constitue plutôt un groupe apparenté aux insectes. Constatation moins surprenante : les crustacés primitifs appelés Remipedia constituent un groupe parent non éteint proche des insectes.
Bernhard Misof, un co-auteur de l'étude, souligne que l'exploration de l'ensemble de données génétiques que son équipe a généré n'est pas terminée car les chercheurs n'ont pas utilisé toutes les données de séquence “Le principal objectif était de trier entre ce qui était plausible et robuste et ce qui devait être rejeté”, explique Bernhard Misof. “Nous fournissons un arbre qui constitue l'épine dorsale de l'arbre phylogénétique des insectes”.
Le schéma ci-dessous représente une vision modernisée de l'arbre phylogénétique de l'immense classe des insectes. À l'extérieur du cercle, sont notés les 29 ordres d'insectes (en noir) et leurs plus proches parents (en blanc), puis, à l'intérieur, les 146 genres les plus importants. Les branches de l'arbre et leurs nœuds montrent les relations entre ces groupes en remontant le fil de l'évolution jusqu'au centre, où se trouve l'ancêtre commun à tous ces arthropodes.
Pour zoomer, cliquer sur le schéma
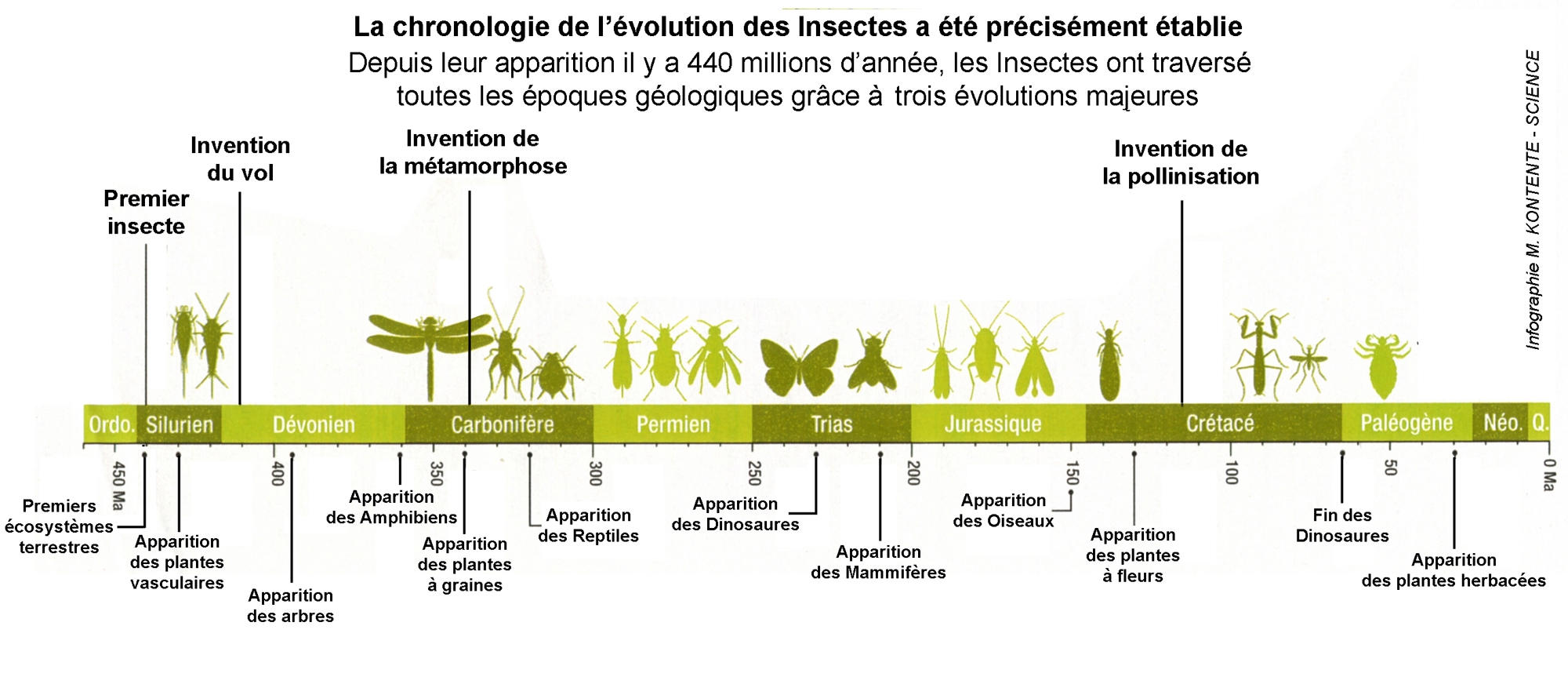
Les recoupements moléculaires permettent d'estimer l'apparition des hexapodes, et donc des premiers ancêtres des insectes, à 479 millions d'années (Ordovicien), alors que se formaient à peine les premiers écosystèmes terrestres. Les insectes eux-mêmes ont émergé, il y a 440 millions d'années. Et leur succès ne s'est jamais démenti. Car quand on parle de 6 extinctions massives des animaux, eux n'en ont connu que... 3 moyennes ! Un tour de force qui repose sur trois inventions, désormais mieux datées.
TROIS INVENTIONS MIEUX DATÉES
- Le vol d'abord, que les aïeux des libellules sont les premiers à maîtriser il y a 406 millions d'années (Dévonien), alors que les écosystèmes commencent à se développer.
- La métamorphose ensuite, qui, à l'instar de l'asticot devenant mouche, bouleverse la morphologie de l'animal à maturité. Elle serait primitivement apparue il y a 3 millions d'années (Carbonifère) chez les holométaboles, pour vraiment se répandre au Crétacé.
- Enfin, dernier coup de génie : la pollinisation et la coévolution avec les plantes à fleurs qui vont accompagner l'explosion des hyménoptères (abeilles), diptères (mouches) et lépidoptères (papillons) au Crétacé.
Le nouveau tracé du parcours évolutif des insectes permet aussi de mieux cerner leurs ancêtres communs. Éphémères et libellules auraient ainsi eu un seul et même parent il y a 360 millions d'années. Quant aux poux et autres parasites, la polémique est relancée : ils ne seraient pas nés, il y a 150 millions, mais depuis 50 millions d'années.
Les chercheurs ne comptent pas s'arrêter là. Avec encore plusieurs millions d'espèces à| découvrir, ce nouvel arbre sera un outil incontournable.
Sources :
É. Rauscher (2015).- Insectes, leur folle diversité enfin mise en ordre Science & Vie, n° 1170 mars 2015, pp. 84-93) article illustré de magnifiques clichés en microscopique électronique à balayage de différents insectes. http://www.science-et-vie.com
À découvrir : les collections du MNHN.
B. Misof et al., “ Phylogenomics resolves the timing and pattern of insect evolution,” Science, 346:763-67, 2014.
À consulter :
> Une présentation de leur travail par plusieurs chercheurs chez The Scientist :
http://www.the-scientist.com/?articles.view/articleNo/413...
> Le site du projet 1Kite « 1000 génomes Insectes Transcriptome Evolution » :
Site sur lequel on peut aussi trouver une vidéo qui retrace l’évolution des premiers insectes sur terre (en anglais) :
http://www.1kite.org/news.html
> Un dossier en ligne de l’université Berkeley qui permet de replacer l’évolution des insectes dans celle de la planète (en anglais) :
http://nature.berkeley.edu/~oboyski67/download/UCSC.pdf
À lire
> Le Guide critique de l’évolution (éditions Belin, 576 pages, 36 €), sous la direction de Guillaume Lecointre (MNHN) :
http://www.editions-belin.com/ewb_pages/f/fiche-article-g...p
> Et son compagnon indispensable, La Classification phylogénétique du vivant, toujours aux éditions Belin (560 pages, 43 €), par Guillaume Lecointre et Hervé Le Guyader :
http://www.editions-belin.com/ewb_pages/f/fiche-article-l...
> En France, le Muséum national d’histoire naturelle possède une belle collection d’insectes, y compris fossiles :
http://www.mnhn.fr/fr/collections/ensembles-collections/p...
18:14 Publié dans Biologie, Insectes, Invertébrés | Tags : insectes, phylogénie, phylogenèse, arbre phylogénétique | Lien permanent | Commentaires (0) | Facebook | | |
25/11/2014
La Grande Galerie de l'Évolution fête ses vingt ans


10:59 Publié dans Actualité des Sciences, Biologie | Lien permanent | Commentaires (0) | Facebook | | |
09/05/2014
Courbet et les phanères
COURBET et les PHANÈRES
par Claude Roland Marchand
Professeur honoraire des Universités
C’est en examinant de près avec l’œil du biologiste que j’ai découvert, dans quelques tableaux de Gustave Courbet des détails singuliers qui méritent des commentaires sinon des interrogations.
Sur plusieurs tableaux connus j’ai relevé une représentation très subjective des phanères portés par les mammifères et les oiseaux. Je précise qu’on appelle phanères ces productions cutanées des vertébrés : poils, cheveux, plumes, cornes, bois, griffes, sabots… Je ne m’attarderai pas sur les poils pubiens, d’autres l’ont fait avant moi et mieux que moi ; encore que, la polémique sur L’Origine du Monde ne soit pas close en ce début d’année 2014 ! Courbet n’en finit pas de défrayer la chronique, de se dérober et de dissimuler ses mystères… Il convoque, il provoque et va jusqu’à choquer sciemment.
Le cerf dans « l’Hallali »
Sans m’attarder sur la symbolique du tableau, et sur l’artificialité de la mise en scène (saison, région, cavalier, chiens, neige…), j’ai porté mon regard sur les bois du cervidé. Bel animal à l’agonie orné de bois magnifiques. Tous les andouillers sont bien orientés, sauf un : il est courbé vers l’avant, sur la ramure gauche. J’ai observé de nombreux trophées, dans les musées, sur les gravures mais aucun ne présente une telle disposition des cors. Le cerf de ce tableau est une exception, une rareté. Et cet andouiller ainsi orienté représente peut-être une menace, un avertissement « Ne me touchez pas ! » Courbet ne l’a pas peint sans arrière-pensée. On sait qu’il s’est projeté dans ses mises en scène ; et ce cerf à l’agonie est une métaphore du lynchage médiatique auquel il a été confronté lors de plusieurs salons. Mais l’animal cache une arme redoutable qui peut blesser si on s’approche trop près de sa tête.

Ces schémas A et B nous montrent de beaux bois où les premiers andouillers sont tous courbés vers l’arrière. D’après Beaumont et Cassier (1987)

Ce que Courbet nous propose
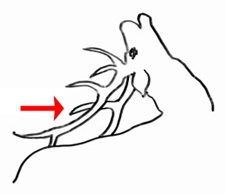
Ne serait-ce pas mieux ainsi ? (croquis C. R. Marchand)

Un cerf du Musée de la chasse me donne raison ! © C. R. Marchand
Le chevreuil dans « la Curée »
De toute évidence, l'animal peint par Courbet n’est pas un cervidé : il s'agit d'un bovidé, en l'occurrence d'une antilope. Un cervidé perd ses bois (c’est de l’os) et les renouvelle chaque année. Le bovidé (comme la vache) a des cornes (en kératine) creuses et pérennes.
La légende du tableau de Courbet précise : « La Curée. Chasse au chevreuil dans les forêts du Grand Jura ». Pourquoi a-til fait cette annonce, lui, le chasseur, qui a abattu de nombreux chevreuils ? Où s’est-il procuré cette antilope, qu’il a pendue dans son atelier parisien ? Chez son boucher ? Au Jardin des Plantes ? Mystère… En fait, il ne s’est pas trompé ; il veut nous tromper. Et il souhaite qu’on lui fasse des objections. Bovidé ou cervidé, peu importe. C’est l’animal mort qui retient son attention, et force la nôtre. Ce qui est surprenant c’est que cette bizarrerie, ce contre-sens zoologique, n’ait pas été plus souvent relevé et commenté.
Décidément, ce tableau composite (en cinq morceaux !), étrange, nous interpelle et nous met mal à l’aise. Le chasseur songe, le cor sonne, les chiens se repaissent de sang, la forêt abrite la mort. Cette scène insolite : il n’y a qu’un chasseur ; c’est une chasse à courre achevée par une scène singulière, incongrue pour des chasseurs jurassiens chevronnés. La curée était rare en Franche-Comté et elle l’est toujours, sauf si on a vraiment imposé et respecté le protocole de la vénerie. Où Courbet est-il allé chercher cette scène ? À qui s’adresse-t-il ? À des Parisiens ou à des Francs-Comtois ?



Les mouettes (1867)
Ce tableau peint lors de son séjour en Normandie, présente une fille qui a un peu les traits de Jo l’Irlandaise, et qui transporte trois volatiles marins appelés « Mouettes ». Ce que nous contestons pour de nombreuses raisons.
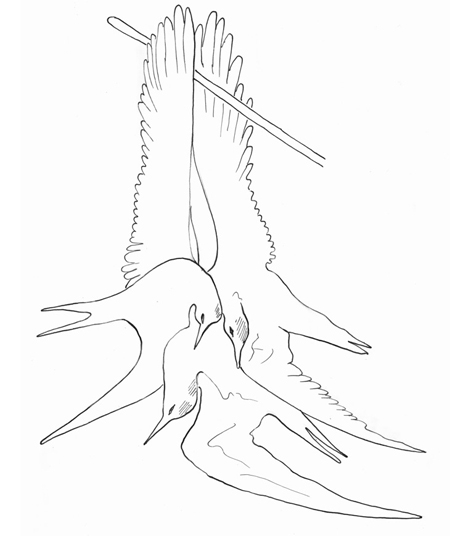
Voici « Les Mouettes » dans « La fille aux Mouettes » 1867.
(croquis C. R. Marchand sur calque)
Examinons les plumes de ces oiseaux marins : leurs queues et leurs ailes falciformes ne sont pas des queues de mouettes, mais sont plutôt des queues de Sternes (caugek ?).

Comparaison des queues de Mouette (à gauche) et de Sterne (à droite)(croquis Marchand)
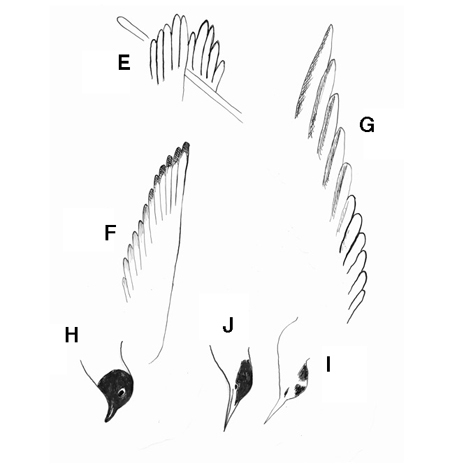
E : les extrémités « digitées » des ailes du tableau de Courbet. F : aile d’une vraie Mouette. G : aile d’une vraie Sterne. H : tête d’une Mouette en été. I : tête de la Mouette du tableau. J : tête d’une vraie Sterne. (croquis de C. R. Marchand)
Courbet le réaliste, ne traduit pas la réalité. Ou ne veut pas l’exprimer. Sont-ce les oiseaux qui importent dans ce tableau, ou est-ce la porteuse des trophées ? Oiseaux libres fauchés dans leur vol et pendus à un bâton, exprimant un rêve brisé ? Une fille volage qui afficherait ostensiblement son tableau de chasse ? Et cette sorte d’imprécation verticale des deux ailes disposées comme deux mains qui saisissent le bâton ?
On se perd en conjectures. Mais ce tableau n’est pas innocent : le peintre nous convoque et pourrait bien être lui-même, l’une de ces fausses mouettes mortellement et irréversiblement blessée.
Une étrange chevelure
Si l’on regarde attentivement l’une des versions du portrait de Jo, la Belle Irlandaise, on peut deviner, dans la chevelure tissée par la main droite, le visage de quelqu’un, ou de quelqu’une. Illusion volontaire, ou hasard du coup de pinceau. Je pose la question.
Courbet a-t-il voulu exprimer ce qu’une main de femme peut emmêler ou démêler ? Est-ce lui, ou un concurrent dans la conquête de la dame ? Est-on avant ou après une défaite amoureuse ? Mystère…


Courbet serait-il lui-même un phanère ?
Ma question peut sembler provocatrice, péjorative, mais je persiste à penser que « Le Chêne de Flagey » héberge, contient, absorbe Courbet.
Examinons l’ombre blanche à droite du tronc. Faisons une copie réduite de l’artiste à Ste-Pélagie, ou de sa photo en pied. Faisons-la glisser sur le tronc ; si la réduction est réussie on voit que la silhouette adossée du peintre se pose exactement à la place des taches de lumière. (cf montage ci-dessous).
Non seulement Courbet y affiche son enracinement, mais il est lui-même l’écorce du chêne. Il est en quelque sorte une émanation du cambium, du liber (liberté ?), l’équivalent d’un phanère végétal si l’on veut bien oser l’analogie jusqu’au bout.

Courbet n’est-il pas lui-même et le double, le fantôme de Vercingétorix ?
Bien ancré dans le sol, accolé au tronc, encore vivant ou déjà mort ?

Même attitude dans La Curée. Mêmes intentions ?
Conclusions
Je pense que cet exposé partial ne sera pas du goût des admirateurs du peintre d’Ornans. J’ai conscience des faiblesses, des limites de mes remarques et de l’audace qui m’a animé pour oser les écrire.
C’est Courbet qui l’a voulu. Et il serait ravi de savoir qu’on découvre une ou deux de ses intentions cachées (j’en aurais d’autres à mentionner…), ou qu’on lui fasse exprimer des sentiments qu’il n’avait peut-être jamais éprouvés ou jamais mis en scène.
Il n’y a pas de tableau banal et muet chez Courbet. Il y parle sans cesse de lui, de ses doutes, de ses désirs, de ses fantasmes parfois.
Moi, en tant que biologiste, j’ai mis ma loupe sur des bizarreries et j’aimerais bien que quelqu’un m’éclaire. Je me sentirais moins seul et moins présomptueux.
Merci Gustave de m’avoir occupé l’esprit souvent et de m’avoir fait douter…
On ne touche pas à Courbet dit-on ; moi, il me touche et ces quelques lignes ont pour prétention de le prouver.
Claude-Roland MARCHAND
Serre les Sapins le 7 mai 2014.
16:04 Publié dans Art et Nature, Biologie | Tags : gustave courbet, cerf, chevreuil, antilope, bois de cerf, bopis de chevreuil, corne d'antilope, artiste peintre | Lien permanent | Commentaires (0) | Facebook | | |
26/12/2013
Cancer : Bactéries intestinales en renfort de la chimiothérapie
Quand les bactéries intestinales viennent en renfort de la chimiothérapie
(Article mis à jour le 5 septembre 2014)
Appelé autrefois "flore microbienne", le microbiote est l'ensemble des microbes (bactéries et champignons essentiellement) qui peuplent notre corps. Ces micro-organismes vivent avec nous, logés à la surface de la peau, dans les cavités buccale et vaginale, et surtout dans notre intestin : celui-ci contient 100 000 milliards de bactéries - soit 10 fois le nombre de nos propres cellules - et leur masse avoisine les deux kilogrammes chez l'adulte !
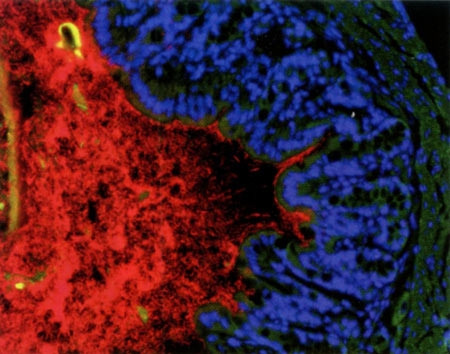
Les bactéries du microbiote apparaissent en rouge
contre la paroi du colon (bleu et vert) © Institut Pasteur
Ce microbiote intestinal est aujourd'hui considéré comme un véritable organe, avec une activité métabolique égale à celle du foie car les espèces bactériennes qui le composent exercent des fonctions cruciales pour notre santé comme l’élimination des substances étrangères à l’organisme (et potentiellement toxiques) ou le maintien à distance de pathogènes qui nous contaminent. Elles assurent également la dégradation des aliments ingérés pour une meilleure absorption intestinale et un métabolisme optimal. Ces milliards de bactéries colonisent l’intestin dès la naissance et jouent un rôle-clef dans la maturation des défenses immunitaires.
Les espèces bactériennes qui composent le microbiote intestinal diffèrent toutefois d'un individu à l'autre et la présence ou l'absence de telle ou telle bactérie semble influencer la survenue de certaines maladies ou au contraire nous protéger. Il nous permet de digérer les sucres complexes et les fibres, intervient dans la biosynthèse des vitamines ou encore dans la détoxification de certaines substances de l'alimentation, et il forme une barrière protectrice contre les pathogènes. Il comprend environ 1000 espèces de bactéries et sa composition varie d'un individu à l'autre. Chacun d'entre nous a donc son propre microbiote intestinal, constitué dans les premières années de la vie à partir des bactéries de notre environnement : seul un tiers des bactéries intestinales est commun à tous.
Ce microbiote intestinal joue un rôle crucial dans le développement du système immunitaire et, sans doute, dans une large gamme de maladies : l'obésité, la résistance à l'insuline et diverses maladies inflammatoires de l'intestin (maladie de Crohn, rectocolite hémorragique...) peuvent résulter d'un déséquilibre du microbiote intestinal. Plus étonnant : il pourrait intervenir dans certaines pathologies de l'humeur comme la dépression. De plus en plus d'études montrent en effet que nos bactéries intestinales envoient des signaux au cerveau et semblent ainsi capables d'influencer le comportement.
Une recherche menée conjointement par des chercheurs de Gustave Roussy, de l'Inserm, de l’Institut Pasteur et de l'Inra a permis une découverte assez étonnante sur la façon dont les traitements de chimiothérapie anticancéreuse agissent plus efficacement grâce à l'aide du microbiote intestinal. Les chercheurs viennent en effet de démontrer que l’efficacité d'une des molécules les plus utilisées en chimiothérapie, repose en partie sur sa capacité à entraîner le passage de certaines bactéries de la flore intestinale vers la circulation sanguine et les ganglions. Une fois dans les ganglions lymphatiques, ces bactéries stimulent de nouvelles défenses immunitaires qui iront aider l'organisme à combattre encore mieux la tumeur cancéreuse.

Arbre phylogénétique bactérien sur fond de muqueuse du côlon d'une souris. La taille des cercles renseigne sur l'importance des populations bactériennes du microbiote. Les points rouges correspondent aux bactéries renforçant l'action d'une immunothérapie anticancéreuse, les verts à celles inhibant la réponse à une chimiothérapie.
Dans le domaine du cancer, l’équipe française* dirigée par le Pr Laurence Zitvogel, à l’Institut Gustave Roussy et directrice de l'Unité Inserm 1015 "Immunologie des tumeurs et immunothérapie", en collaboration étroite avec l’Institut Pasteur (Dr Ivo Gomperts Boneca, Unité "Biologie et génétique de la paroi bactérienne") et des chercheurs de l’INRA (Dr Patricia Lepage et Dr Joël Doré, Unité Micalis "Microbiologie de l'Alimentation au service de la Santé"), vient d'apporter la preuve que la flore intestinale stimule les réponses immunitaires d'un individu pour combattre un cancer lors d'une chimiothérapie.
*Équipe française dirigée par le Pr Laurence Zitvogel, à l'Institut Gustave-Roussy, directrice de l'Unité Inserm 1015 "Immunologie des tumeurs et immunothérapie", en collaboration étroite avec l'Institut Pasteur (Dr Ivo Gomperts Boneca, Unité "Biologie et génétique de la paroi bactérienne") et des chercheurs de l'INRA (Dr Patricia Lepage et Dr Joël Doré), Unité "Microbiologie de l'Alimentation au Service de la Santé Humaine".
Le cyclophosphamide est l’un des médicaments les plus utilisés en chimiothérapie. Comme tout traitement, il entraîne cependant des effets secondaires (inflammation des muqueuses etc.) et perturbe l'équilibre normal du microbiote intestinal. Chez la souris, l'équipe a démontré que le cyclophosphamide modifie la composition du microbiote dans l'intestin grêle et induit la translocation de certaines espèces de bactéries Gram-positives dans les organes lymphoïdes secondaires. Là, ces bactéries stimulent la production d'un sous-ensemble spécifique de "pathogène" T helper(17)" pTH17 cellules mémoire et des réponses immunitaires Th1. La synergie entre le microbiote et le cyclophosphamide débute lorsque le médicament facilite le passage à travers la barrière intestinale des bactéries Gram+ qui vont se retrouver dans la circulation sanguine et les ganglions lymphatiques. Ces bactéries sont considérées comme néfastes et l'organisme déclenche une réponse immunitaire. Une fois dans les ganglions lymphatiques, elles stimulent la production de globules blancs pTH17 qui vont agir comme de nouvelles défenses immunitaires et aider l'organisme à lutter contre la tumeur cancéreuse. "Cette réaction en chaîne, effet secondaire du traitement, va s'avérer en réalité très utile" explique Laurence Zitvogel. "De façon surprenante, la réponse immunitaire dirigée contre ces bactéries va aider le patient à lutter encore mieux contre sa tumeur en stimulant de nouvelles défenses immunitaires."
En détails, l'immunisation anti-bactérienne aboutit au recrutement de lymphocytes effecteurs différents de ceux mobilisés par la chimiothérapie. Leur rôle consiste à aider les lymphocytes anti-tumoraux à endiguer la croissance de tumeurs.
A contrario, des souris dépourvues de tout germe intestinal ou préalablement traitées par des antibiotiques dirigés contre les bactéries Gram+ se révèlent incapables de produire les précieuses cellules pTH17 antitumorales. De plus, leur tumeur est devenue résistante au cyclophosphamide. La situation est toutefois réversible : lorsque les sourisreçoivent une perfusion intraveineuse de pTH17, le médicament retrouve son efficacité antitumorale.
Les chercheurs suggèrent également que certains antibiotiques utilisés au cours d’une chimiothérapie pourraient détruire ces bactéries Gram+ et annuler ainsi leur effet bénéfique. "Maintenant que ces bactéries bénéfiques potentialisant la réponse immunitaire anti-tumorale ont été identifiées, on devrait réussir rapidement à en fournir plus à l'organisme, notamment via des pro- ou pré-biotiques et/ou une alimentation spécifique" conclut la chercheuse.
Ces travaux ont bénéficié du soutien de la Ligue nationale contre le cancer, de l'Institut national du cancer (INCa) (SIRIC SOCRATES) et du LABEX Onco-Immunologie.
Remarque :
Le cyclophosphamide n'est pas le seul traitement à profiter d'un microbiote intestinal performant. Le même numéro de Science fait ainsi part du travail d'une équipe américaine qui aboutit à des conclusions très proches : certains antibiotiques diminueraient l'action de la chimiothérapie.
Parallèlement au travail des chercheurs français, Elizabeth Pennisi, Noriho Lida et al, de l'Institut national du cancer (Frederick, États-Unis), montrent que le microbiote intestinal renforce les effets d'autres traitements anticancéreux, à savoir une immunothérapie ou une chimiothérapie par oxaliplatine (un autre médicament anticancéreux). Ici aussi, comme dans l'étude française, l'efficacité du traitement antitumoral a chuté drastiquement chez des souris débarrassées de leur flore bactérienne intestinale.
Même si ces deux études ont été menées sur la souris, il est vraisemblable que leurs résultats puissent être un jour applicables à l'être humain. Ils suggèrent en tout cas que certains antibiotiques pourraient diminuer l'efficacité d'une chimiothérapie antitumorale mais également qu'une supplémentation en pro- ou pré-biotiques, voire une alimentation spécifique, pourrait renforcer l'action du traitement.
Sources :
Laurence Zitvogel et al (2013). - The Intestinal Microbiota Modulates the Anticancer Immune Effects of Cyclophosphamide, Science22 November 2013, vol 342 n° 6161 pp. 971-976.
Elizabeth Pennisi et al (2013). - Cancer Therapies Use a Little Help From Microbial Friends, Science 22 November 2013 Vol. 342 n° 6161 p. 921.
Marc Gozlan (2013). - La flore intestinale en renfort de la chimiothérapie, Sciences et Avenir, janvier 2014 n° 803, p. 70.
Qu'est-ce que le microbiote ? Lettre de l'Institut Pasteur, septembre 2014 n° 86 p. 9.
11:55 Publié dans Actualité des Sciences, Biologie, Médecine | Tags : cancer, chimiothérapie, microbiote intestinal, flore intestinale | Lien permanent | Commentaires (0) | Facebook | | |
04/11/2013
Stage de permaculture à Froidevaux
Stage de permaculture à Froidevaux
Ce stage de permaculture est co-organisé par une nouvelle association créée pour l'occasion (en fait le potager d'une curieuse avec quelques amis) et Bernard Alonso, permaculteur canadien qui est déjà venu à Besançon en 2012.
La permaculture est un ensemble de pratiques et de modes de pensée visant à créer une production agricole soutenable, très économe en énergie (travail manuel et mécanique, carburant...) et respectueuse des êtres vivants et de leurs relations réciproques. Elle vise à créer un écosystème productif en nourriture ainsi qu'en d'autres ressources utiles, tout en laissant à la nature "sauvage" le plus de place possible.
Elle utilise des notions d'écologie, de paysagisme, d'agriculture biologique, de biomimétisme, d'éthique, de philosophie et de pédologie. La permaculture invite à mettre ces aspects théoriques en relation avec les observations réalisées sur le terrain.
La base de la permaculture n'est pas uniquement d'analyser les éléments constitutifs d'un système individuellement, mais aussi de prendre en compte leurs interactions, dans le but de produire une compréhension de l'écosystème dans l'optique d'une utilisation par l'Homme.



08:13 Publié dans Biologie, Botanique, Environnement-Écologie, Nature & philosophie, Nature et santé | Lien permanent | Commentaires (0) | Facebook | | |
03/11/2013
Vidéo : l'ourse Hvala et ses deux oursons
Une vidéo exceptionnelle de l'ourse Hvala et ses deux oursons de l'année
C'est probablement la plus belle vidéo depuis les derniers lâchers d'ours en 2006 dans les Pyrénées : l'ourse Hvala et ses deux oursons nés en 2013 filmés en plein jour près d'un arbre équipé d'une caméra automatique du réseau Ours brun.
Des images exceptionnelles qui répondent d'elles-mêmes aux démagogues qui présentent le retour de l'ours comme un échec ...
08:01 Publié dans Biologie, Environnement-Écologie, Mammifères | Lien permanent | Commentaires (0) | Facebook | | |
02/11/2013
Le premier cerveau éprouvette
Le premier cerveau éprouvette
par Aude Rambaud
(Science & Vie n° 1154, novembre 2013, pp. 92-95)
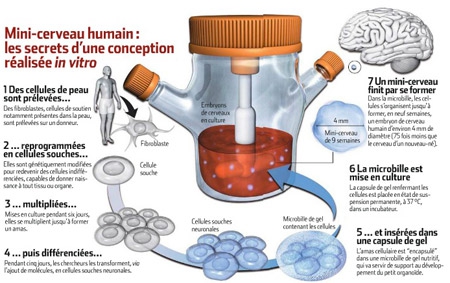
À première vue, ce n'est qu'un petit amas de cellules. Une sphère d'à peine 4 millimètres de diamètre, maintenue en suspension dans un liquide transparent. Rien, dans l'apparente insignifiance de cet objet biologique, ne laisse présager son importance. Et pourtant ! Prévenu de sa véritable nature, l'observateur ressent une indicible émotion. Car ce qu'il a sous les yeux, flottant dans une petite boîte, n'est ni plus ni moins que le tout premier embryon de cerveau humain cultivé en laboratoire. Une réplique vivante du futur organe de la pensée, comme il se présente chez un fœtus d'un peu plus de deux mois. Un choc ! Un choc et... une incroyable prouesse, que l'on doit aux chercheurs de l'Institut de biotechnologie moléculaire de Vienne (Autriche).

Encapsulés dans des microbilles de gel nutritif (à droite) placées en suspension dans un incubateur (à gauche), des amas de cellules souches ont donné naissance à un embryon de cerveau humain (ci-dessous, en coupe)
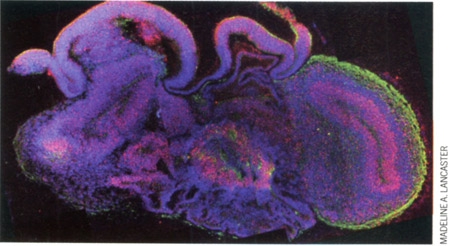
À l'origine de cet exploit, il y a les dernières découvertes en matière de cellules souches, ces cellules capables de donner naissance à tout type de tissu ou d'organe, et dont les potentialités ne laissent d'impressionner. Ainsi, à partir de quelques cellules de peau prélevées sur un donneur et génétiquement "reprogrammées" au stade de cellules souches, les chercheurs ont obtenu, deux mois et demi plus tard, la réplique quasi parfaite du cerveau du donneur... tel qu'il devait être aux prémices de son développement embryonnaire, après neuf semaines de gestation. "C'est un travail magnifique", commente, admiratif, Étienne Hirsch, directeur adjoint du Centre de recherche de l'institut du cerveau et de la moelle épinière (Paris). Pas de doute, la "fabrication" in vitro de l'organe le plus complexe du corps humain, même à un stade primaire, repousse les frontières du possible. En même temps qu'elle interroge les limites de la culture d'organes.
Des matrices pour organes
Le pari fou des chercheurs autrichiens n'était pourtant pas gagné d'avance. Certes, depuis le milieu des années 2000, les biologistes maîtrisent de mieux en mieux les cellules souches. Ils savent guider leur transformation, par des techniques de mise en culture, pour qu'elles donnent toutes les sortes de lignées cellulaires qui composent un être humain : cellules cardiaques, musculaires, pancréatiques... ou même neuronales. Mais sans support "physique", les cellules souches se bornent à former de simples tapis biologiques. Dans le même temps, d'autres équipes ont donc mis au point des matrices, sortes de charpentes pour culture cellulaire en trois dimensions, qui ont déjà permis de faire pousser in vitro des copies plus ou moins parfaites d'organes, comme la vessie, les vaisseaux sanguins ou le foie.
Presque comme un vrai !
Étant donné sa complexité, le cerveau restait toutefois hors de portée. Principale difficulté ? Il est constitué de neurones aux formes différentes et de cellules de soutien (cellules gliales) qui s'imbriquent les unes dans les autres. Un puzzle d'une précision telle qu'il semblait impossible à reproduire. "Les précédentes tentatives avaient abouti à des ensembles de neurones de même type. Cette fois, nous avons un véritable petit organe en développement, présentant tous les aspects d'un cerveau in vivo, malgré une organisation un peu anarchique ", estimait Jürgen Knoblich, auteur des travaux, lors de la présentation des résultats.
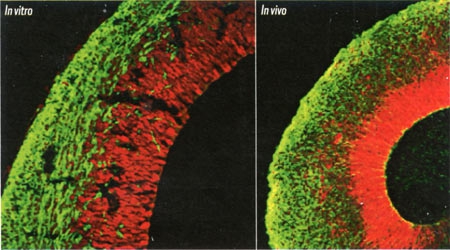
Le cortex embryonnaire a une structure identique in vivo (à droite) et in vitro (à gauche), même si l'organisation des cellules y est anarchique
Le secret ? Il ne réside pas dans le matériel biologique utilisé. En effet, les chercheurs autrichiens s'en sont tenus aux cellules souches classiquement obtenues. Lesquelles, mises en culture, se sont notamment différenciées en neuroectoderme, le tissu à l'origine du cerveau. Jusqu'ici, rien de vraiment neuf. Puis, les chercheurs ont découpé des fragments de ce tissu, qu'ils ont encapsulés dans des microbilles de gel nutritif contenant toutes les substances nécessaires à la croissance et au développement des cellules en culture. Voilà l'une des premières astuces. "L'utilisation de microbilles à la place d'une couche de gel au fond d'une boîte a amélioré l'adhérence entre les cellules", explique Mathilde Girard, de l'Institut des cellules souches I-Stem d'Évry.
Ensuite, et c'est là qu'est la véritable clé du succès, ils ont utilisé un bioréacteur tournant, une sorte d'incubateur qui a placé les microbilles en état de suspension permanente. "C'est la grande nouveauté du dispositif, s'enthousiasme la chercheuse. Les billes en suspension contournent le problème de la gravité. Lorsque les cellules commencent à former un amas au sein de la capsule de gel, elles ne se déforment pas et ne retombent pas comme dans une boîte de Pétri. En outre, cette mobilité permet d'améliorer l'oxygénation des cellules. "Autrement dit, c'est en se rapprochant des forces mécaniques exercées sur ces cellules durant une grossesse que les chercheurs ont triomphé.
Frottements, gravité, mouvement... Le résultat de cette modification de l'environnement est étonnant. Dans leurs microbilles, les cellules se multiplient, se différencient et s'organisent de façon totalement autonome. Comme si les cellules avaient en mémoire le plan à respecter pour aboutir à un cerveau, avec des régions cérébrales bien distinctes : cavités remplies de liquide qui donneront les ventricules et le liquide céphalo-rachidien, couches internes du cortex cérébral avec neurones différenciés, plexus choroïdes produisant le liquide céphalo-rachidien, ou encore tissu rétinien chez environ 10 % des embryons de cerveau. Autant de structures présentes in vivo et qui montrent même des traces d'activité électrique identiques... "Certaines étapes du développement sont tout à fait semblables à ce qui se passe in vivo. Néanmoins, l'organisation spatiale au sein de ces régions, anarchique, ne reproduit pas l'architecture réelle d'un cerveau", tempère Jean Bernard Manent, chercheur à l'Institut de neurobiologie de la Méditerranée, à Marseille.
Un développement bloqué à neuf semaines
Cette première étape franchie, le cerveau n'a cependant pas continué à se développer. Pourquoi ? Les chercheurs ont bien essayé de pousser plus loin l'expérience, en laissant leur culture dans l'incubateur au-delà de neuf semaines... Mais si le mini-cerveau a pu survivre pendant plusieurs mois, l'absence de circulation sanguine, source d'oxygène et de nutriments, a empêché sa croissance. "Sans système vasculaire, le développement s'arrête rapidement. Or, il semble impossible de le reconstituer, car il faudrait pour cela reproduire la totalité de l'embryon in vitro ", explique Mathilde Girard.
Ces travaux n'en intéressent pas moins les médecins, qui espèrent utiliser ces cerveaux embryonnaires pour tester la toxicité de certains médicaments lors des premiers mois de la grossesse, ou étudier des pathologies liées au développement cérébral à partir des cellules d'un individu malade. Ils nous font pénétrer plus loin que jamais dans les coulisses de l'être humain en formation. L'homme parviendra-t-il un jour à créer en laboratoire un véritable cerveau humain parvenu à son stade adulte de développement ? L'avenir le dira. Mais l'idée, à la fois fascinante et effrayante, ne relève plus tout à fait du domaine de la science-fiction.
L'informatique aussi modèle le cerveau
In vitro ou in silico ? Modéliser le cerveau humain pour tester sur lui de futurs médicaments, c'est aussi l'un des objectifs poursuivis par le Human Brain Project. Cet autre projet fou compte reproduire, à l'aide de simulations informatiques, le fonctionnement cérébral d'ici dix ans (voir Science & Vie n° 1145, p. 44). Près de 80 instituts de recherche européens et internationaux regroupent et analysent actuellement toutes les données disponibles : biologiques, cliniques, issues de l'imagerie, etc. À terme, cette cartographie devrait permettre de reconstituer les circuits neuronaux, de modéliser certaines maladies neurologiques ou encore de tester des molécules. Si la majeure partie des recherches concerne le cerveau adulte, il est prévu d'ajouter au projet des données sur le développement cérébral, grâce à des clichés de cerveaux d'enfants prématurés ou plus âgés.
Quatre dates importantes sur ce sujet
1998 : découverte des cellules souches embryonnaires (CSE) chez l'Homme.
2006 : création de cellules souches induites, dites iPS, à partir de cellules de peau.
2009 : culture de neurones fonctionnels en laboratoire à partir de CSE.
2013 : fabrication du premier embryon de cerveau humain à partir de cellules iPS.
11:29 Publié dans Actualité des Sciences, Biologie, Médecine | Lien permanent | Commentaires (0) | Facebook | | |
10/10/2013
Les cellules souches pluripotentes induites (iPS)
Les cellules souches pluripotentes induites (iPS)
par Hervé Ratel
journaliste scientifique
(Sciences et Avenir n° 794, avril 2013, pp. 66-68)
(dernière mise à jour : 28 février 2014)
Le sigle iPS désigne les cellules souches pluripotentes induites, c'est-à-dire des cellules adultes rajeunies au stade embryonnaire. Un stade où elles sont capables de donner n'importe quel type de cellule de l'organisme (pluripotence). Les iPS ouvrent des perspectives majeures dans le traitement de nombreuses maladies incurables. Les premiers essais vont commencer au Japon.
Rajeunir les cellules, c'est la prouesse annoncée par ce simple sigle : iPS. Ce terme désigne les cellules souches pluripotentes induites, soit des cellules adultes reprogrammées jusqu'au stade primitif de cellules embryonnaires. Pour nombre de scientifiques, la révolution annoncée est comparable à celle des vaccins ou des antibiotiques en leur temps. Mise sous les projecteurs en octobre 2012 avec le prix Nobel décerné à leur découvreur, le Japonais Shinya Yamanaka (voir Sciences et Avenir n° 789, novembre 2012), ces cellules iPS sont en effet susceptibles de rendre d'immenses services dans le domaine des greffes de tissus, de faciliter la mise au point de médicaments pour des maladies génétiques et d'élucider certains mécanismes du cancer. Un comité d'éthique japonais a d'ailleurs donné son feu vert pour de premiers essais cliniques qui devraient débuter dès le mois d'avril 2013.
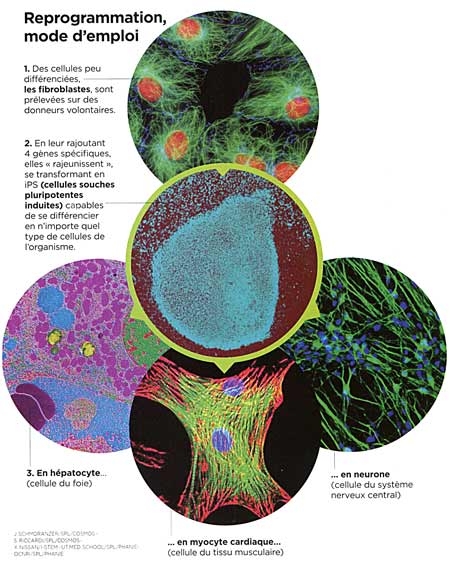
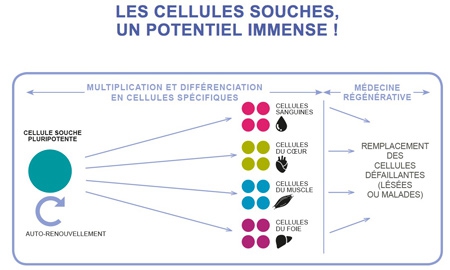
Diagramme © Institut Pasteur
Dans la course mondiale qui démarre, la France est bien placée. "Elle pourrait prendre la tête d'un consortium chargé de piloter un projet de banque européenne de cellules iPS. Celui-ci sera finalisé d'ici à la fm de cette année", pronostique Annelise Bennaceur-Griscelli (Inserm), qui coordonne la plate-forme nationale de cellules souches Ingestem, rassemblant plusieurs laboratoires de l'Hexagone.
Pourtant, les chercheurs français reviennent de loin ! En cause : la loi de bioéthique de 1994 - révisée en 2004 - qui interdit toute recherche sur les cellules souches embryonnaires humaines (CSEh). Ces dernières - qui ne sont pas des iPS - nécessitent en effet d'avoir directement recours à des embryons surnuméraires issus de fécondations in vitro. « Pour obtenir une dérogation les autorisant à travailler sur les CSEh, les chercheurs devaient remplir des documents faisant plusieurs dizaines de pages, raconte Frank Yates, enseignant-chercheur à l'école Sup'Biotech de Paris. Il fallait par exemple décrire à l'avance - exercice impossible et absurde les progrès médicaux attendus des expériences envisagées... Certains baissaient les bras ou s'exilaient à l'étranger devant ce cauchemar bureaucratique.» D'autres pays comme le Royaume-Uni, ont une réglementation moins restrictive, plus en phase avec la recherche fondamentale.

Les scientifiques abasourdis par ces résultats
La découverte des iPS en 2006 est donc venue bouleverser la donne puisque ces cellules ne nécessitent comme matériel de départ que des cellules adultes déjà différenciées, prélevées sur des donneurs consentants. Lors de leur découverte annoncée en 2006 dans la revue Cell c'est pourtant le scepticisme qui avait dominé parmi la communauté scientifique. « J'ai eu la chance d'assister à la première conférence du PrYamanaka sur le sujet, se remémore Frank Yates. Quand il a présente ses résultats, tout le monde était abasourdi. Parvenir à reprogrammer une cellule paraissait totalement improbable. » Improbable parce que cette découverte réduisait à néant l'un des dogmes majeurs de la biologie : une cellule différenciée, comme une cellule musculaire, ne pouvait pas se transformer en une cellule d'un autre type, une cellule hépatique par exemple. Pourtant, il n'a pas fallu longtemps pour que les résultats du chercheur japonais soient reproduits et confirmés par d'autres équipes de biologistes cellulaires à travers le monde. Les cellules iPS étaient bel et bien une réalité.
Pour comprendre, il faut revenir à l'intuition de génie de Shinya Yamanaka : le chercheur a voulu savoir ce qui se passerait si on transférait des gènes actifs uniquement au stade embryonnaire, dans une cellule différenciée adulte. Pour mener l'expérience, il a, avec son équipe, identifié dans un premier temps 24 gènes de ce type, avant de restreindre la sélection à 4 d'entre eux (Oct4, Sox2, Klf4 et c-Myc). Et ça a marché ! Les cellules adultes ont "remonté le temps" jusqu'à redevenir des cellules indifférenciées. Mais cette recette est évidemment bien plus délicate à mettre en œuvre. Et elle ne fonctionne pas à tous les coups : « Dans le meilleur des cas, le taux de conversion de cellules adultes en iPSn'excède pas 5 % », précise Jean-Marc Lemaitre (Inserm, Montpellier). De très nombreux obstacles restent donc à franchir avant de trouver des applications directes aux iPS.
Un travail de culture astreignant fait à la main
La première difficulté consiste à obtenir ces fameuses iPS en quantités suffisantes. Car développer une lignée, c'est un peu comme réussir un soufflé : les ingrédients sont peu nombreux, mais rien n'est possible sans un bon tour de main, du doigté et de la patience. Une fois une iPS obtenue au bout de quelques jours, il faut en effet absolument éviter qu'elle ne se différencie à nouveau et ne redevienne une cellule adulte. Comment ? En veillant en permanence à séparer les iPS nouvellement créées du reste de leurs consœurs encore à l'état différencié dans la boîte de Pétri, ces récipients où l'on cultive les cellules en laboratoire. Un travail astreignant qui, aujourd'hui, se fait encore manuellement. Trois semaines de culture sont nécessaires pour obtenir une colonie et il faut plusieurs mois avant que cette colonie compte quelques milliers de spécimens. « De plus, les cellules iPS sont très sensibles et capricieuses. Il faut les surveiller 24 h/24 », explique Lina El Kassar. Tous les trois mois, cette biologiste de l'I-Stem au Génopole d'Evry, dans l'Essonne, accueille ainsi des chercheurs dans son atelier pour leur transmettre son savoir-faire. À l'issue du stage, chacun repart avec ses propres lignées d'iPS sur lesquelles il peut poursuivre ses travaux de recherche. Une initiative fructueuse qui a permis de former rapidement à la culture des iPS quantité de biologistes français, contribuant au bon positionnement de la France.
Autre difficulté : les iPS souffrent de défauts épigénétiques. En effet, la cure de jouvence qu'elles subissent ne parvient pas à remettre parfaitement tous les compteurs biologiques à zéro. Ainsi, les iPS - à l'inverse des CSEh, les cellules souches embryonnaires humaines - conservent de petites « étiquettes » à la surface de leurs gènes indiquant toujours leur signature de cellules adultes. Autrement dit, vues sous l'angle épigénétique, les iPS n'ont que l'apparence de cellules embryonnaires. En profondeur, elles demeurent de "vieilles" cellules. « Que ce soit pour unefuturemédecine régénératrice ou comme modèle in vitro d'une pathologie, ces désordres épigénétiques doivent êtrepris en compte, explique Marc Peschanski, directeur scientifique de l'I-Stem. Il faudra vérifier au préalable qu'ils ne constituent pas une nuisance.» Car le risque que des iPS souffrant de tels défauts deviennent cancéreuses n'est pas à écarter.
Obtenir des cellules souches pluripotentes par balnéation acide ? (janvier 2014) (Sciences et Avenir n° 805, mars 2014, p. 23)
Plonger des cellules dans un bain légèrement acide est-il de nature à les stresser suffisamment pour les transformer en cellules souches comme le proclame l'équipe de Haruku Obokata, du centre Riken, à Kobe ?
Depuis la publication fin janvier dans les colonnes de Nature du travail des Japonais, des critiques n'ont pas tardé à apparaître sur Internet, et plusieurs équipes dans le monde ont tenté de reproduire ces résultats, pour l'instant, la moisson n'est pas fameuse.
Selon le décompte du site Ipscell, sur dix tentatives, une seule s'est révélée fructueuse, et encore avec une production très modeste. « J'aurais tendance à classer ce résultat des Japonais dans les artefacts de culture, avoue Frank Yates, enseignant-chercheur à Sup'Biotech. Intuitivement, nous sommes beaucoup à penser que s'il était aussi simple d'obtenir des cellules souches, nous y serions arrivés bien avant par accident... »
Irrégularités méthodologiques, fraude délibérée, erreurs de manipulation ou travail scientifique révolutionnaire de nature à changer la face des biotechnologies ? La réponse définitive viendra sous peu, à la fin des investigations scientifiques de rigueur.
Des champs d'application très larges
Étudier une maladie.
En prélevant des cellules sur un malade, les iPS permettent de suivre le développement d'une cellule affectée d'une mutation. Ce qui était impossible avec les CSEh, car il faut disposer d'embryons porteurs de la pathologie.
Mieux comprendre le cancer.
En suivant le devenir d'une cellule à partir de son stade embryonnaire, les chercheurs espèrent élucider les mécanismes conduisant aux processus tumoraux et notamment les métastases.
Concevoir de nouveaux médicaments.
Les iPS doivent permettre à terme détester facilement l'effet de molécules potentiellement intéressantes pour bon nombre de maladies. Cette recherche est actuellement au point mort.
Élaborer une médecine régénératrice.
C'est l'objectif ultime : pouvoir remplacer des neurones, des cellules musculaires, un épiderme, etc. à partir de cellules suffisamment proches du système immunitaire du malade. Voir également sur ce même blog les articles : cellules souches et rajeunissement, des cellules souches dans la rétine, régénération de la rétine humaine, élaboration d'une rétine humaine en culture à partir de cellules souches, obtenir des neurones à l'aide de fibroblastes, créer des dents biologiques grâce à des cellules souches,
L'industrie pharmaceutique n'est pas prête à investir
Les espoirs se tournent plutôt vers la constitution de banques de cellules iPS qui seraient immunologiquement compatibles avec la majeure partie d'une population donnée : Européens, Asiatiques, etc. Soit des banques ethniques qui risquent de faire grincer bien des dents... D'autant que les firmes pharmaceutiques ne semblent pas décidées à participer à la constitution de telles banques. « Culturellement, les industriels de la pharmacie sont des chimistes, pas des biologistes, justifie Marc Peschanski. Ils ont encore du mal à travailler avec du matériel vivant. » Un avis partagé par Laurence Daheron, responsable de la plate-forme iPS/CSEh à l'université Harvard (Cambridge. États-Unis) : « L'industrie pharmaceutique commence tout juste à entrevoir le potentiel des iPS. Elle est loin d'être prête à s'investir dans la thérapie cellulaire. Il n'y a aucun changement radical à attendre de sa part pour les années à venir. Elle se contentera d'utiliser les iPS pour rester des médicaments classiques, rien de plus. »
Pourtant, tout porte à croire que le jour où la recherche publique aura prouvé le potentiel thérapeutique des iPS, la demande de millions de patients en attente d'un traitement sera suffisamment pressante pour forcer la main des industriels. Il reste juste à espérer que ces derniers auront alors la capacité de prendre le train en marche...
S'affranchir des manipulations fastidieuses in vitro en produisant in vivo des cellules souches pluripotentes induites (iPS)
Dans un article de Nature paru le 11 septembre 2013, des chercheurs espagnols sont parvenus à reprogrammer « in vivo » des cellules adultes de souris. Cette technique permet donc de s'affranchir des fastidieuses et coûteuses mises en culture des cellules adultes !
Le premier défi pour les chercheurs du Centre national de recherche sur le cancer (CNIO, Madrid) était de reproduire l'expérience de Yamanaka chez un être vivant. Ils ont choisi la souris comme organisme modèle. En utilisant des techniques de manipulation génétique, les chercheurs ont créé des souris dans lesquelles quatre gènes de Yamanaka pouvaient être activés à volonté. Lorsque ces gènes sont activés, les chercheurs ont observé que les cellules adultes étaient capables de régresser dans leur développement évolutif pour devenir des cellules souches embryonnaires dans plusieurs tissus et organes
En outre, l'équipe de Manuel Serrano a ainsi obtenu des cellules souches de meilleure qualité que celles produites in vitro. C'est un résultat spectaculaire : faire en sorte que des souris fabriquent des cellules souches pluripotentes induites, capables de fournir quasiment n'importe quelle cellule spécialisée de l'organisme. Autrement dit, qu'au sein d'un animal vivant des cellules adultes redeviennent des cellules souches embryonnaires ! Reprogrammer in vivo des cellules est désormais possible. La démonstration chez l'animal de la faisabilité d'un tel concept pourrait bouleverser la pratique de la future médecine régénératrice.
Les chercheurs espagnols ont ainsi franchi une nouvelle étape, en obtenant les mêmes résultats que Yamanaka, mais cette fois au sein du même organisme, chez la souris, sans avoir besoin de passer à travers dans des boîtes de culture in vitro. La génération de ces cellules dans un organisme apporte cette technologie encore plus proche de la médecine régénérative.
L'étape du laboratoire sera ainsi supprimée. Jusqu'à présent, on imaginait que pour greffer ces cellules iPS à un malade souffrant de diabète insuline-dépendant, d'insuffisance cardiaque ou de la maladie de Parkinson, il fallait au préalable les produire en passant par des manipulations sophistiquées et coûteuses en laboratoire. Le travail réalisé par l'équipe de Manuel Serrano bouleverse ce protocole : il sera possible d'induire la production de ces précieuses cellules souches au sein même du tissu ou de l'organe à réparer sans passer par l'étape du laboratoire. Une fois ces iPS produites par le patient, elles seraient amenées à se différencier en un type cellulaire particulier, en cellule du pancréas, du cœur, en neurone, par exemple.
Comme indiqué au début de cet article, pour reprogrammer des cellules adultes (de peau notamment) en cellules souches, il faut y introduire quatre gènes clés (Oct4, Sox2, Kfl4 et c-Myc). Ceux-ci produisent quatre facteurs de croissance jouant un rôle dans les premiers temps du développement embryonnaire. Les cellules adultes ainsi reprogrammées, dénommées iPS, sont capables de « remonter le temps » jusqu'au stade embryonnaire.
Cette technique de production, mise au point en 2006, a d'ailleurs valu le prix Nobel de médecine en 2012 au Japonais Shinya Yamanaka. Les chercheurs espagnols sont allés plus loin : l'organisme de leurs souris transgéniques fabrique lui-même ces quatre facteurs de croissance. Pour activer les quatre gènes, ils ont fait boire aux rongeurs de l'eau contenant de faibles doses d'un antibiotique : la doxycycline. Résultat : au bout de deux semaines et demie, les rongeurs ont développé des tumeurs (tératomes,) issues de cellules souches pluripotentes et contenant plusieurs types cellulaires adultes. Preuve manifeste pour les chercheurs que des cellules adultes de ces rongeurs avaient bien été reprogrammées en iPS avant qu'elles ne se transforment, à leur tour, et en divers endroits, en plusieurs types cellulaires. Par ailleurs, l'analyse de tissus de l'estomac, de l'intestin, du pancréas et des reins des rongeurs a montré la présence disséminée d'une reprogrammation cellulaire in vivo. De même, les chercheurs ont détecté la présence d'iPS dans le sang des souris qui avaient été reprogrammées pour en fabriquer.
Ces cellules iPS sont d'une qualité supérieure
Une autre surprise attendait les chercheurs : les cellules iPS produites chez ces souris et obtenues directement à partir de l'intérieur de l'organisme, ont une plus grande capacité de différenciation que ceux obtenus par culture in vitro. Plus précisément,ces ceklules ont les caractéristiques des cellules totipotentes, un état primitif jamais obtenu dans un laboratoire.
En effet, outre leur capacité à se transformer en n'importe quel type de cellule d'un embryon, elles ont pu se différencier en placenta. Ce résultat étonnant fait dire aux chercheurs que ces iPS obtenues in vivo sont à un stade encore plus précoce que les iPS produites en laboratoire. Il est évidemment hors de question d'utiliser une procédure semblable chez l'Homme, dans la mesure où les cellules reprogrammées risqueraient de former des tumeurs. En revanche, une stratégie de régénération in vivo pourrait, dans l'avenir, consister à administrer au malade, localement et pendant un temps limité, des gènes synthétiques codant les quatre facteurs de croissance identifiés par le prix Nobel Shinya Yamanaka.
D'autres pistes sont à l'étude comme administrer, au sein d'un organe ou d'un tissu malade, un cocktail de petites molécules capables d'activer le programme génétique de production de cellules iPS ou utiliser des virus rendus inoffensifs pour délivrer les quatre fameux gènes clés.
María Abad, le principal auteur de l'article et chercheur dans le groupe de Serrano, a déclaré: "Ce changement de direction dans le développement n'a jamais été observé dans la nature, nous avons démontré que nous pouvons également obtenir des cellules souches embryonnaires dans les organismes adultes et pas seulement en. le laboratoire ".
"Nous pouvons maintenant commencer à penser à des méthodes pour induire la régénération localement et de manière transitoire pour un tissu endommagé particulier.", précise Manuel Serrano.
Les cellules souches obtenues chez la souris montrent également des caractéristiques totipotentes jamais produites dans un laboratoire, équivalentes à celles présentes dans les embryons humains au stade de 72 heures de développement, quand ils sont composés de seulement 16 cellules.
En comparaison avec les cellules obtenues avec la technique développée par Yamanaka, les cellules souches obtenues par le CNIO représentent donc un état encore embryonnaire plus précoce, avec une plus grande capacité de différenciation.
Les auteurs ont même été capables d'induire la formation de structures pseudo-embryonnaires dans les cavités thoracique et abdominale des souris. Ces pseudo-embryons affichaient les trois couches typiques des embryons (ectoderme, mésoderme et endoderme), et des structures extra-embryonnaires tels que la membrane vitelline et même l'ébauche de cellules sanguines.
"Ces données nous disent que nos cellules souches sont beaucoup plus polyvalentes que les cellules iPS obtenues in vitro par Yamanaka, qui génèrent les différentes couches de l'embryon mais jamais les annexes embryonnaires, comme le placenta", a déclaré Manuel Serrano.
Les auteurs soulignent que les applications thérapeutiques possibles de leurs travaux restent lointaines, mais ils admettent que, sans doute, cela pourrait signifier une réorientation de la recherche sur les cellules souches, la médecine régénérative et l'ingénierie tissulaire.
"Nos cellules souches survivent aussi en culture, permettant ainsi de les manipuler dans un laboratoire. L'étape suivante est d'étudier si ces nouvelles cellules souches sont capables de produire efficacement différents tissus tels que ceux du pancréas, du foie ou du rein. ", a déclaré Abad.
Sources :
- María Abad, Lluc Mosteiro, Cristina Pantoja,Marta Cañamero, Teresa Rayon,Inmaculada Ors, Osvaldo Graña, Diego Megías, Orlando Domínguez,Dolores Martínez, Miguel Manzanares,Sagrario Ortega & Manuel Serrano. (2013). Reprogramming in vivo produces teratomas and iPS cells with totipotency features Nature : 10.1038/nature12586.
- Marc Gozian (2013) Des souris vivantes ont produit des cellules souches (Sciences et Avenir, n° 800, octobre 2013, pp. 78-79).
Voir également dans ce blog, l'article : Les-potentialités infinies des cellules souches.
16:50 Publié dans Biologie, Médecine | Tags : cellules souches, cellules souches adultes induites, ips, cseh, rétine, cancer | Lien permanent | Commentaires (0) | Facebook | | |
03/10/2013
Les bactéries font et défont les espèces
 Les bactéries font et défont les espèces
Les bactéries font et défont les espèces
par Émilie Rauscher
(Science & Vie n° 1152, septembre 2013, pp. 106-111)
Les bactéries logées à l'intérieur de tout organisme vivant, y compris celles de la flore intestinale, influencent son évolution… Au point, parfois, de créer de nouvelles espèces. L'hypothèse avait été formulée il y a quelque temps déjà... En 1927, le microbiologiste américain Ivan Wallin notait :
"C'est une proposition pour le moins étonnante de dire que les bactéries, communément associées à la maladie, pourraient en fait être un facteur fondamental dans l'apparition de nouvelles espèces..."
Tellement étonnante, d'ailleurs, qu'elle fut rapidement oubliée, d'autant que les arguments pour la démontrer n'existaient pas... Ou plutôt pas encore. Car aujourd'hui, alors qu'ils mesurent encore à peine à quel point les bactéries sont partout – des abysses à nos cellules – ; alors qu'ils commencent à réaliser l'importance des relations ultimes et durables (symbioses) qu'elles ont tissées avec tout ce qui vit ; alors qu'ils découvrent enfin tout ce dont elles sont capables... finalement, les scientifiques en viennent à se dire qu'il y avait peut-être du vrai dans cette vieille idée folle !
Selon Bordenstein, professeur de biologie à l'université Vanderbilt (Nashville, EtatsUnis), lui, en est convaincu depuis plus de dix ans déjà : "Une nouvelle espèce pourrait bien apparaître, non seulement parce que ses gènes mutent au sein de ses propres cellules, mais aussi parce que les communautés microbiennes qu'elle héberge (dans ses intestins, etc.) changent ! " Et il a désormais de quoi étayer cette hypothèse, qui non seulement renouvelle notre compréhension de la complexe mécanique de l'évolution, mais fait des bactéries l'un des maîtres, aussi majeur que méconnu, du destin des espèces...
NOUS SOMMES TOUS DES ÉCOSYSTÈMES
Inspiré par Ivan Wallin, mais aussi par d'autres prédécesseurs visionnaires comme la biologiste américaine Lynn Margulis, qui avait perçu dès les années 1970 l'importance de la symbiose, Seth Bordenstein a patiemment réuni cas ambigus et phénomènes étranges, comme autant d'indices de l'influence des bactéries. Il mène des recherches, compile et dissèque les travaux dans lesquels des micro-organismes semblent être à l'origine d'une nouvelle espèce... Une gageure tant ce phénomène, la "spéciation", est discuté par les scientifiques : ses influences sont innombrables et ses causes multiples et difficiles à distinguer de ses conséquences.
Déjà se pose la question de ce qu'est une espèce. Cela semble simple si on regarde la chose d'un œil presbyte : de loin, les espèces se distinguent bien. De près, beaucoup moins. La faute à l'évolution et à la sélection naturelle qui poussent le vivant à changer en permanence, si bien qu'il est difficile de faire la part des choses quand une espèce se transforme ou que deux populations proches semblent s'hybrider. Par conséquent, depuis deux cent cinquante ans qu'ils travaillent dessus, les naturalistes, puis les biologistes ne se sont toujours pas accordés sur une définition claire... Retenons juste que des individus appartiennent à des espèces différentes s'ils ne peuvent pas se reproduire ensemble. Les raisons d'une telle barrière ? Soit l'acte est impossible entre eux, soit leurs hybrides sont condamnés ou stériles. Dans tous les cas, les ponts ont été coupés entre les partenaires et entre leurs espèces respectives — qui pouvaient n'en faire qu'une au départ. Incapables d'échanger des gènes, elles vont suivre chacune leur chemin évolutif et multiplier les différences. C'est ainsi qu'en partant d'un cousin de Velociraptor, on se retrouve quelques millions d'années plus tard avec des canards et des autruches. D'où viennent ces différences ? Comme le rappelle Seth Bordenstein, "les biologistes considèrent que l'apparition de nouvelles espèces est principalement dirigée par les changements qui se produisent dans nos gènes. Nous, nous démontrons que pour comprendre de façon globale comment se passe la spéciation, il faut certes considérer notre génome, mais aussi celui des micro-organismes qui sont en nous !" Pour le biologiste américain, la "cause première" de la séparation des espèces peut être, aussi, microbienne. Rappelons-nous que nous sommes de vrais écosystèmes, peuplés de nuées de bactéries. Tout comme deux populations appartenant à une même espèce peuvent être séparées physiquement par une montagne, ou génétiquement par un bouleversement chromosomique, elles peuvent l'être par ces mini-résidents !
Le chercheur a regroupé une collection inédite de cas où des bactéries ont sorti une artillerie lourde capable d'imposer cette ségrégation...
UN STRATAGÈME PARFAIT
Wolbachia est, à ce titre, exemplaire : cette bactérie infecte près de 60 % des arthropodes, l'immense groupe des insectes, acariens, araignées et crustacés. Parasite incontournable, elle s'insinue dans les cellules de son hôte et colonise la descendance des femelles. Pour favoriser sa transmission, elle réalise des prodiges.
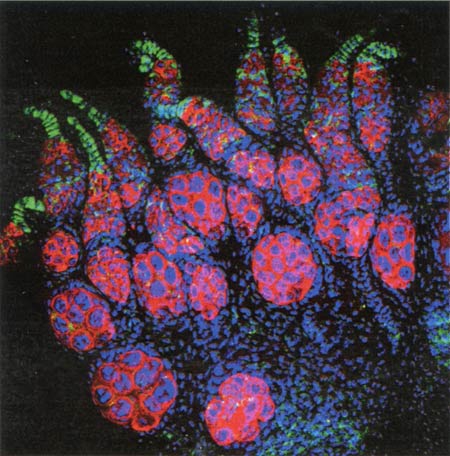
En infectant les cellules souches des organes reproducteurs de la Drosophile, la bactérie Wolbachia (en vert) modifie en sa faveur l'évolution de son hôte.
Ainsi, elle est capable d'imposer à ses hôtes une "spéciation asexuelle" : les femelles qu'elle infecte produisent sous sa houlette des ovules qui se développeront sans être fécondés par les mâles (qui deviennent ainsi inutiles). Chez des guêpes ou des acariens par exemple, on voit ainsi apparaître des lignées entièrement féminines et autonomes. "Ce cas est emblématique car immédiat, pointe Olivier Duron, spécialiste de l'évolution des symbioses à l'université de Montpellier 2. Dès l'arrivée de la bactérie, on a un isolement reproducteur, c'est-à-dire que l'individu infecté ne peut plus se reproduire avec les autres." Et qui dit isolement dit début de divergence entre les populations séparées.
Wolbachia a une seconde technique : elle sait influer sur les possibilités de reproduction de son hôte. Seules les femelles qu'elle infecte peuvent se croiser avec tous les mâles, transmettant ainsi le parasite à sa descendance. Une femelle non infectée est limitée aux mâles également non infectés, sous peine de voir ses œufs mourir. En quelques générations, près de 100 % de la population est parasitée. Ce procédé prend toute sa force quand on sait qu'il existe plusieurs types incompatibles de Wolbachia : ainsi, dans une même espèce d'arthropodes, on peut voir apparaître des groupes infectés par l'une ou l'autre souche bactérienne qui ne pourront plus se croiser !
DES ESPÈCES NAÎTRAIENT D'UN SIMPLE CHANGEMENT DE LEUR FLORE INTESTINALE
Preuve supplémentaire de la responsabilité de la bactérie : l'utilisation d'antibiotiques, en la tuant, fait tomber les barrières qu'elle dressait entre ses hôtes... À moins de s'y prendre trop tard, alors que l'évolution a déjà créé des lignées isolées. Séparées à l'origine par la bactérie, ces populations sont devenues trop différentes pour se croiser, même après sa disparition.
Ces exemples, Seth Bordenstein en connaissait l'importance, mais aussi les limites. Après tout, qu'un parasite niché au cœur même des cellules de son hôte ait un tel effet sur lui n'est sans doute pas si surprenant. Et si Wolbachia et ses consœurs sont courantes chez les arthropodes, on n'en connaît guère chez les mammifères, pour ne citer qu'eux. Ce qui limite leur impact à des groupes animaux, certes nombreux, mais bien particuliers...
Sauf que les bactéries n'ont pas besoin de s'incruster dans les cellules de leur hôte pour bouleverser sa vie. Partout dans le vivant, on observe des symbioses au cours desquelles elles se contentent de s'installer dans certains organes ! Pour rester chez les arthropodes, elles leur permettent de diversifier leur nourriture ou de s'adapter à un autre milieu, et donc de coloniser des zones interdites à leurs congénères. Grâce à elles, le termite mange du bois, le puceron conquiert de nouveaux végétaux, etc. "Puisqu'elle ouvre l'accès à une niche particulière, et donc facilite la divergence avec la population d'origine, la symbiose soutient la spéciation", souligne Olivier Duron. Voilà qui démultiplie les cas possibles !
Escherichia coli
Pourtant, Seth Bordenstein avait l'ambition d'aller encore plus loin. Pour ce défi, il reçoit en 2008 le soutien de son collègue Robert Brucker. Dans leur dernière étude, tout juste publiée, les deux biologistes soutiennent cette fois que même des bactéries banales, non symbiotiques, telles celles qui pullulent dans nos intestins, peuvent provoquer la spéciation ! "Ces dernières années, il est devenu clair que chaque animal avait des hôtes bactériens spécifiques, dont les gènes réunis (le microbiome) sont impliqués dans sa digestion comme dans son bon développement cérébral, rappelle le chercheur. Mais personne n'avait prouvé leur rôle dans la spéciation..."
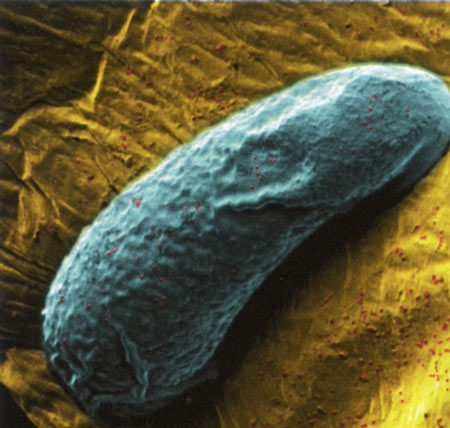
Embryon hybride de Nasonia
Leurs bactéries créent une divergence entre les guêpes qui les empêche de se reproduire. Un traitement antibiotique a mis en valeur l'influence de la flore bactérienne de 3 guêpes du genre Nasonia qui les empêche de se reproduire. Sans traitement, leurs embryons hybrides ne survivraient pas, leurs flores étant incompatibles.
Avec l'aide de trois petites guêpes du genre Nasonia, d'une bonne dose d'antibiotiques et de logique, c'est désormais chose faite. Les chercheurs savaient que lorsque ces trois espèces se croisaient, leurs hybrides étaient condamnés. Ils prédirent, en s'inspirant des expériences avec Wolbachia, qu'un traitement antibiotique aurait un effet drastique... Ils ne se sont pas trompés : près de 90 % des hybrides furent sauvés ! L'incompatibilité entre les guêpes venait donc bien des bactéries. Et réinjecter les microbes restaurait leur lourde mortalité précédente. Un scénario qui a un air de déjà-vu... Sauf que, différence fondamentale, les microbes visés étaient, cette fois, simplement ceux de la flore intestinale !
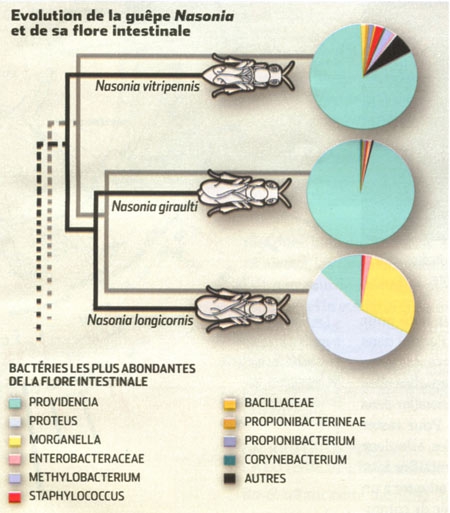
UN JOUEUR DE PLUS À LA LOTERIE DE L'ÉVOLUTION
En étudiant cette dernière de plus près, Seth Bordenstein remarqua d'ailleurs que les hybrides survivant sans traitement étaient ceux qui avaient reçu une flore proche de celle d'un de leurs parents, au lieu d'un mélange chaotique. C'est donc bien cette flore qui détermine qui vit ou meurt et, ce faisant, imperméabilise les frontières entre les espèces concernées. "Cette étude est solide et l'idée qu'elle présente est séduisante. On savait que les bactéries intracellulaires pouvaient jouer sur l'isolement de populations... Mais que les bactéries intestinales en soient également capables est nouveau ! ", s'enthousiasme Olivier Duron. Ainsi, les espèces - et l'homme ne ferait pas exception - pourraient naître et prospérer par le fait de mutations dans leur génome mais aussi, simplement, par des changements dans leur flore intestinale.
Voilà une découverte qui ajoute un joueur particulièrement riche à la grande loterie de l'évolution... Et qui dit partenaire supplémentaire dit nouveaux gènes à prendre en compte : "Notre expérience donne ainsi une bonne raison de regarder 1'hologénome, c'est-à-dire la combinaison de l'ADN de l'hôte et de ses communautés de micro-organismes résidents, souligne Seth Bordenstein. L'objet de la sélection naturelle ne serait donc pas l'individu seul, mais lui et les communautés bactériennes associées." Pour le chercheur, nous étions jusqu'ici passés à côté de la plus grande part de l'information qui constitue chaque être vivant : les bactéries, ces marionnettistes invisibles et inconscients derrière l'évolution des espèces !
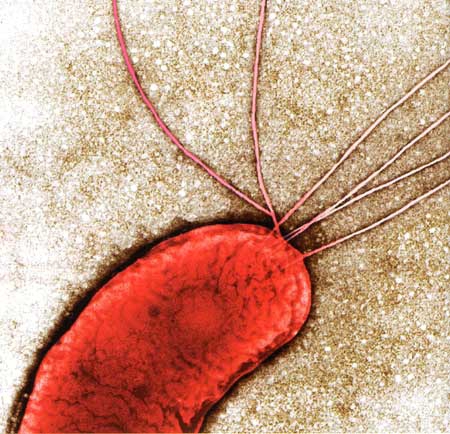
Proteus vulgaris
ET CHEZ L'HOMME ?
Notre part bactérienne est plus importante qu'on ne le croit : pour dix cellules humaines, nous abritons quatorze bactéries et pour dix gènes humains, nos microbes en comptent quatorze. Outre nos quelque vingt milliers de gènes, nous en comptons donc quelque trente milliers d'origine microbienne : deux patrimoines génétiques qui doivent fonctionner "en réseau".
Le rôle des bactéries dans l'évolution des insectes décrypté, on peut se demander ce qu'il en est des mammifères ou des poissons. Seth Bordenstein et Robert Brucker se posent bien sûr la question : "Nous espérons montrer que la flore intestinale et les gènes des microbes qui la composent, le microbiome, sont importants pour la spéciation animale. Une étude récente vient d'ailleurs de montrer que l'Homme et ses plus proches parents chez les primates ont des microbiomes spécifiques à chaque espèce, qui évoluent en même temps que l'ADN nucléaire (dans le noyau des cellules)." Cette coévolution plaide pour un lien fort entre notre génome et celui de notre flore. Les études se multiplient d'ailleurs pour recenser les bactéries que nous hébergeons et mieux comprendre le microbiome riche de plusieurs millions de gènes qu'elles constituent. On sait déjà qu'il existe trois "groupes bactériens" intestinaux, comme il y a des groupes sanguins, et que les perturber a des conséquences sur notre santé.
20:16 Publié dans Biologie, Médecine | Tags : spéciation, génomique, bactéries, flore bactérienne, microbiome | Lien permanent | Commentaires (0) | Facebook | | |
04/03/2013
L'Établissement français du sang (EFS)
L'Établissement f rançais du sang (EFS)
rançais du sang (EFS)
Outre son rôle d'opérateur de la transfusion sanguine, l'Établissement français du sang (EFS) recouvre des activités partant de la recherche fondamentale jusqu'à l'élaboration de protocoles de thérapie et l'administration de soins. Au cœur de cette organisation, le département des activités d'ingénierie cellulaire et tissulaire de l'EFS Bourgogne - Franche-Comté met en œuvre les résultats de la recherche, qui elle-même s'adosse à son expertise et à son savoir-faire, en une sorte de cercle vertueux dédié à l'immunologie de la greffe et du cancer.
Recherche et ingénierie cellulaire : des échanges de bons procédés
Les cellules souches hématopoïétiques sont au centre des activités de l'EFS Bourgogne – Franche-Comté. À l'origine des différentes lignées de cellules sanguines, elles se trouvent chez le sujet adulte dans la moelle osseuse, véritable usine à globules rouges, globules blancs et plaquettes.
Lorsque la production est perturbée, voire rendue stérile par certaines attaques cancéreuses ou les traitements, la thérapie cellulaire relance le mécanisme selon différents protocoles pour tirer le patient de son aplasie. L'autogreffe consiste à prélever dans la moelle osseuse d'un patient des cellules réputées saines à la suite de cycles de chimiothérapie. Conservées par congélation, elles seront utilisées pour pallier le déficit cellulaire qui ne manquera pas de se présenter lors de traitements ultérieurs destinés à détruire les cellules cancéreuses résiduelles, puissants au point d'atteindre aussi les cellules indemnes.
À Besançon, la première autogreffe a été réalisée en 1979 sous l'impulsion du professeur Patrick Hervé. Elle est pratiquée en routine depuis les années 1990. L'autogreffe présente des limites dans les cas de myélomes très agressifs ou de leucémies aiguës, pour lesquels les spécialistes ont recours à l'allogreffe, comme à Besançon depuis 1983.
Cherche cellules souches compatibles
Lors de l'allogreffe, le prélèvement de cellules souches hématopoïétiques est effectué sur un donneur présentant une typologie HLA compatible, de préférence de la famille du patient, à défaut inscrit sur des registres de volontaires en France et dans le monde.
Le facteur temps est ici capital pour la réussite de la greffe. Fabienne Pouthier, responsable médicale du département AICT de l'EFS Bourgogne – Franche-Comté raconte combien chaque cas est unique. « Récemment, nous avons trouvé à Washington un donneur compatible pour un patient suivi à Besançon. Le prélèvement de moelle osseuse opéré aux États-Unis est arrivé le lendemain par transport ultrasécurisé, pour une greffe réalisée le soir même au CHRU Jean Minjoz. »
L'alternative du sang placentaire
Plus récentes en France, avec une réelle généralisation depuis 2005, les greffes de cellules souches hématopoïétiques issues de sang placentaire représentent une alternative lorsque la compatibilité avec les donneurs fait défaut. Si les récentes directives internationales encouragent la création de banques de sang placentaire, Besançon s'est montrée pionnière en France avec la création à l'EFS du premier dispositif de ce type en 1995.
Les atouts majeurs du sang placentaire sont sa disponibilité immédiate et la jeunesse de ses cellules souches hématopoïétiques, caractérisées par un potentiel de régénération optimum, un risque de rejet greffon-hôte moindre et une absence de contamination virale. Leur immaturité implique cependant un temps plus long pour se développer dans l'organisme et vaincre l'aplasie.
Aux côtés des banques de cellules souches hématopoïétiques hébergées sur le site bisontin de l'EFS, figure également une banque de tissus, la seule banque EFS pour tout l'Est de la France. Les tissus, recueillis auprès des donneurs selon une démarche similaire à celle du don d'organes, sont essentiellement des cornées, des vaisseaux sanguins et des valves cardiaques, stockés pour être utilisés lors de greffes futures ou à des fins clé recherche.
Il va sans dire que toutes ces activités sont soumises aux règles les plus strictes de contrôle et de sécurité, tant dans le prélèvement et la conservation des cellules et des tissus que dans l'élaboration des produits à des fins de thérapie ou de recherche.
Ainsi l'EFS Bourgogne - Franche-Comté souhaite obtenir prochainement la labellisation de Centre de production de médicaments de thérapies innovantes préparés ponctuellement (MTI PP) répondant, aux normes européennes les plus récentes et donnant une dynamique nouvelle à une activité et un savoir-faire déjà reconnus.
Contact : Fabienne Pouthier - Département des activités ingénierie cellulaire et tissulaire - EPS Bourgogne - Franche-Comté
Tél. (0033/0) 3 81 61 50 27 - fabienne.pouthier@efs.sante.fr

Recherches pionnières en thérapie génique
II n'est pas étonnant qu'avec une production de vingt millions de cellules à la seconde, notre organisme, à la mécanique pourtant bien rodée, présente quelques défaillances. Des erreurs corrigées par le système immunitaire, qui lui, a parfois du mal à faire face à toutes les sollicitations dont il fait l'objet et peut laisser échapper à sa vigilance des cellules dangereuses, à l'origine de tumeurs.
Une armée de lymphocytes T
Lors du contrôle permanent exercé par le système immunitaire, les lymphocytes T occupent un rôle majeur et se mobilisent en masse lorsqu'il s'agit de combattre des corps étrangers comme les virus, les bactéries ou les cellules cancéreuses. Les recherches s'orientent vers l'utilisation de ces lymphocytes, capables de s'immiscer dans tout l'organisme, pour détruire des cellules tumorales résiduelles difficiles à débusquer, complétant ainsi de manière la plus fine possible les traitements réalisés par chirurgie, radiothérapie ou chimiothérapie. L'allogreffe permet d'une part le remplacement de la moelle osseuse défectueuse d'un patient et d'autre part l'implantation de lymphocytes T suffisamment différents de ses propres cellules pour encore mieux détecter et détruire les cellules nuisibles, persistantes (leucémiques ou lymphomateuses), par un processus dit d'alloréactivité.
Cependant, les lymphocytes T reconnaissent également les cellules saines du patient, risquant de provoquer la maladie du greffon contre l'hôte, mortelle dans environ 30 % des cas. Pour parvenir à un point d'équilibre et éviter de franchir cette frontière ténue entre effet et rejet, les chercheurs de l'équipe de Thérapeutique immuno-moléculaire des cancers (TIM-C) de l'unité mixte de recherche Interaction hôte-greffon / tumeur et ingénierie cellulaire et génique travaillent dans deux directions.
Gène suicide et cible idéale
La première, l'immunothérapie allogénique, consiste à programmer les lymphocytes T pour l'autodestruction, une sécurité qui sera déclenchée si leur action se retourne contre l'organisme hôte. Une fonction rendue possible par manipulation génétique, grâce à l'apport d'un gène « suicide » à activer en cas de complications. L'administration d'un médicament entraîne alors le processus de destruction. Au terme d'essais cliniques qu'ils étaient les premiers à démarrer voilà dix-sept ans, les chercheurs bisontins vont continuer à établir la démonstration de ce concept précurseur, en collaboration avec une équipe américaine du Baylor College of Medicine (Houston), dans un nouveau programme de tests menés sur une population de douze patients à partir de l'automne prochain. Guider le système immunitaire de façon très ciblée contre la tumeur est la deuxième voie explorée, sous le nom d'immunothérapie antitumorale. Il s'agit ici de faire en sorte que les lymphocytes détectent uniquement les cellules tumorales visées grâce à une particularité qu'elles possèdent en propre, et qu'ils dirigent leur action contre elles et elles seules. Dès lors, des lymphocytes produits par clonage cellulaire et moléculaire pourront être injectés en grande quantité et à des moments choisis pour un maximum d'efficacité, sans risque d'effet délétère sur les cellules saines de l'organisme qu'ils ne seront plus à même de reconnaître comme potentiellement dangereuses.
Contact : Christophe Ferrand - Unité mixte de recherche Interaction hôte-greffon / tumeur et ingénierie cellulaire et génique EPS Bourgogne - Franche-Comté / Université de Franche-Comté/ INSERM Tél. (0033/0) 3 81 61 56 15 - christophe.ferrand@efs.sante.fr
Produit anti-inflammatoire à visée thérapeutique
Et si le meilleur anti-inflammatoire était produit par notre propre corps ? Partant du processus biologique du renouvellement cellulaire, au cours duquel les cellules indésirables de l'organisme se détruisent par apoptose, des chercheurs en immunologie ont mis au point un produit thérapeutique capable de combattre l'inflammation.
Cinquante à soixante-dix milliards de cellules sont éliminées chaque jour par notre organisme, et de façon efficace car elles sont potentiellement dangereuses. Pour être actif, ce processus s'opère cependant à l'insu de notre système immunitaire dont il « endort » la vigilance en créant un micro-environnement aux effets immuno-modulateurs puissants.
L'idée est d'utiliser ce micro-environnement pour neutraliser ailleurs et dans d'autres circonstances une réponse inflammatoire incontrôlée de la part du système immunitaire. « Dans cet environnement se trouvent tous les éléments permettant de cibler et maîtriser l'ensemble de la réaction inflammatoire, donc tous les facteurs et cellules impliqués. À l'inverse, les médicaments traditionnels tels que les biothérapies ne ciblent généralement qu'un facteur ou une cellule », explique Sylvain Perruche, chercheur INSERM au laboratoire bisontin Interaction hôte-greffon / tumeur et ingénierie cellulaire et génique.
Ce processus biologique a été copié ex vivo et le micro-environnement généré in vitro. Des tests précliniques ont ensuite confirmé, et de façon spectaculaire, le traitement de souris atteintes d'arthrite rhumatoïde aiguë à un stade avancé. Au terme de dix injections, voire moins selon la concentration du produit, chez une cinquantaine de cas étudiés, la courbe indiquant la flambée arthritique s'inverse radicalement : l'inflammation est totalement jugulée et les fonctions originelles des zones touchées sont restaurées.
Action naturelle, efficacité, polyvalence, facilité de conservation... ce PTC (Produit de thérapie cellulaire) sans cellule verra ses performances protégées par un brevet déposé en janvier dernier sous la houlette de INSERM TRANSFERT. Co-inventeur du brevet, Sylvain Perruche entend poursuivre l'aventure en acquérant le moment venu la licence d'exploitation par le biais d'une start up chargée de la commercialisation du produit.
Contact : Sylvain Perruche - Laboratoire Interaction hôte-greffon / tumeur et ingénierie cellulaire et génique EFS Bourgogne - Franche-Comté / Université de Franche-Comté / INSERM - Tél. (0033/0) 3 81 61 56 15 - sylvain.perruche@inserm.fr
Source :
En Direct, le journal de la recherche et du transfert de l'Arc jurassien n° 247 - mars-avril 2013.
23:11 Publié dans Biologie, Médecine | Lien permanent | Commentaires (0) | Facebook | | |
28/02/2013
Audition et surdités
 L'Oreille,
L'Oreille,
l'Audition
et
les surdités
I. L'audition permet la communication
L'audition est le sens de l'ouïe, c'est-à-dire de la perception des sons.
1. Qu'est-ce que le son ?
Le son est lié à une variation de pression du milieu. Il correspond à la mise en oscillation d'un milieu. Dans le cas de l'homme, il s'agit généralement de l'air, mais l'émission et la conduction sonores peuvent se faire également en milieu liquide ou solide. L'énergie est ensuite transmise, de molécule à molécule, à une vitesse de 340 m.s-1 (dans l'air). Les ondes sonores correspondent à l'alternance de zones de pression élevée et de zones de pression faible. L'amplitude des variations de pression est qualifiée de pression sonore. Elle peut être mesurée à l'aide d'un microphone, qui traduit la variation de pression en une variation de tension électrique.
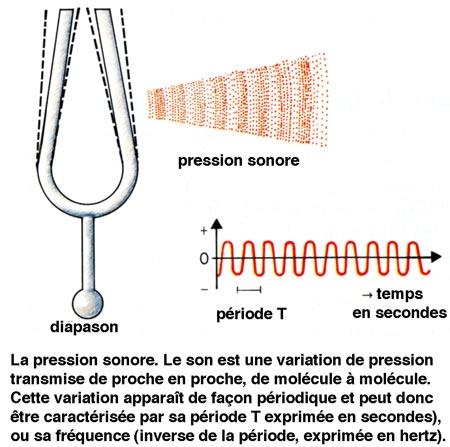
Compte tenu de l'importance de la gamme des niveaux sonores, l'échelle utilisée pour quantifier les pressions sonores est logarithmique. Le niveau de pression de référence PO a été placé légèrement en dessous du seuil de stimulation moyen de l'oreille. Pour une fréquence de 1000 Hz ; il est égal à 2.10-5 N.m-2. Ainsi, le niveau de pression sonore S d'un son donné est égal à :
S = 20 log (P/Po)
L'unité de S est appelée le décibel (dB). À 1 000 Hz, un décibel équivaut à un phone.
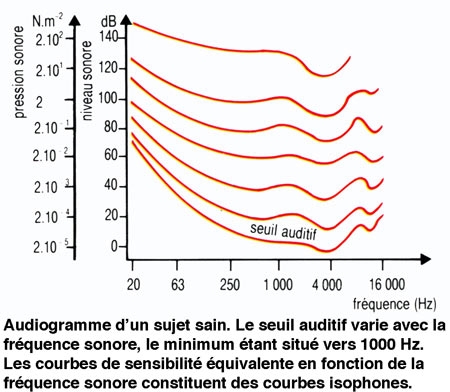
Un second paramètre du son est sa fréquence. Celle-ci est exprimée en cycles par secondes ou herz (Hz). Un son constitué d'une seule fréquence est qualifié de son pur. Cependant, un son est généralement constitué d'un mélange de plusieurs fréquences : une fréquence minimale ou fondamentale et des fréquences multiples de cette fondamentale ou harmoniques. Un son dans lequel toutes les fréquences sont représentées de façon quasi identique est qualifié de bruit blanc.
2. La sensibilité de l'ouïe varie avec la fréquence du son
L'homme est sensible à des vibrations sonores comprises entre 20 Hz et 20 000 Hz (ou 20 kHz). Le seuil d'audition varie en fonction de la fréquence d'un son pur stimulant. Il est minimal pour une fréquence de 1 kHz, et de l'ordre de 4 dB ou 4 phones. Au-delà de ce seuil, la sensibilité varie également en fonction de la fréquence du son stimulant. Ce phénomène peut être représenté sous forme de courbes isophones. À partir d'une valeur d'environ 130 phones, une sensation douloureuse remplace la sensation auditive. De tels sons peuvent endommager l'appareil auditif. Les ondes sonores émises au cours du langage parlé correspondent aux zones de meilleure sensibilité auditive.
3. Des sons graves et des sons aigus
La fréquence sonore est généralement décrite sous forme de hauteur d'un son. Un son grave est un son de basse fréquence, et un son aigu un son de haute fréquence. Dans la musique occidentale, la fréquence sonore est organisée en octaves. Un accroissement de la valeur tonale d'une octave s'obtient en doublant la fréquence d'un son.
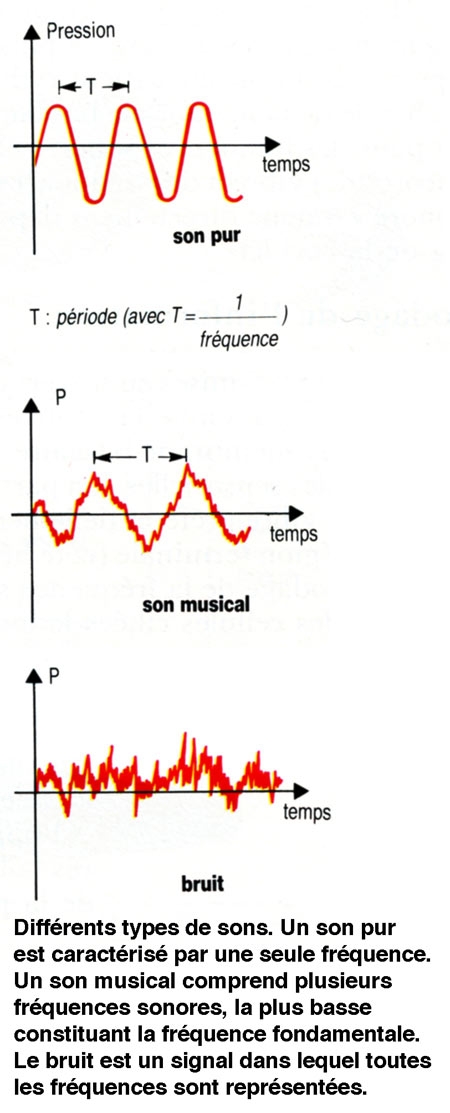
4. Deux oreilles pour faciliter l'orientation auditive dans l'espace
Une écoute à l'aide des deux oreilles permet à un sujet de situer la source sonore dans l'espace. Le son étant conduit à vitesse finie, il ne parvient en même temps au niveau des deux oreilles que si rémission sonore a lieu dans l'axe de la tête. Dans les autres cas, l'onde sonore atteint les deux oreilles avec un décalage temporel qui permet au sujet de détecter la position de l'émetteur sonore.
II. Anatomie de l'oreille et physiologie de l'audition
1. Les sons sont canalisés avant de frapper l'oreille interne
L'oreille représente l'organe de l'audition. En fait, ce que nous avons coutume d'appeler l'oreille n'est que sa partie apparente. L'oreille se compose en réalité de trois parties : externe, moyenne, et interne.. Le conduit entre l'oreille externe et l'oreille moyenne est séparé par une fine membrane : le tympan.
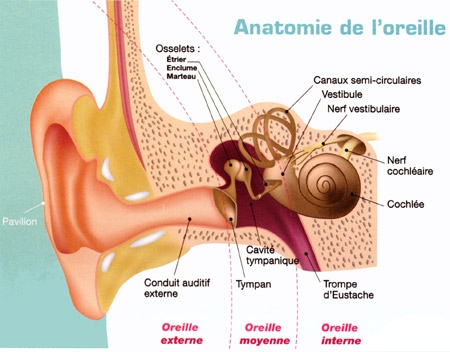
Sa partie visible est l'oreille externe qu'on appelle pavillon, nous sert d'antenne acoustique et de capteur. Les sons pénètrent dans le conduit auditif externe.
Les ondes sonores mettent ensuite en vibration le tympan qui obture l'oreille moyenne, située dans une cavité osseuse du crâne. Dans cette cavité, une chaîne de trois osselets, le marteau, l'enclume et 1'étrier, permet de transmettre les vibrations du tympan vers une structure qui sépare oreille moyenne et oreille interne : la fenêtre ovale. Les trois osselets vont servir d'amplificateurs pour compenser la perte d'énergie liée au passage de l'onde sonore du milieu aérien au milieu liquidien de la cochlée dans l'oreille interne. Saisissant les déplacements du tympan, le marteau transmet son énergie à l'enclume, qui la communique à l'étrier.
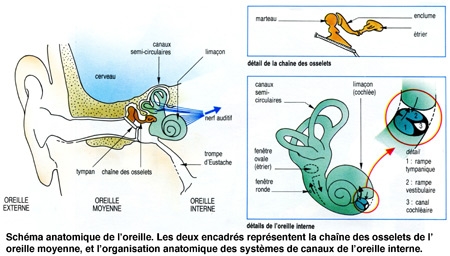
Pour égaliser les pressions de part et d'autres du tympan un conduit, la trompe d'Eustache relie la cavité de l'oreille moyenne au pharynx, une fonction très utile surtout lors du décollage d'un avion par exemple.
L'étrier (le plus petit os du corps humain) est en contact avec la membrane obturant la fenêtre ovale, point d'entrée dans l'oreille interne où se trouve l'organe de l'audition : la cochlée ou limaçon, sorte de long cône composé de trois tubes ou rampes caractérisés par un enroulement hélicoïdal et remplis de liquide.
III. L'oreille interne permet de coder les fréquences sonores
1. La membrane basilaire vibre en fonction de la fréquence sonore
Les vibrations, qui atteignent la fenêtre ovale par l'intermédiaire de la chaîne des osselets, sont transmises au travers des différentes rampes, jusqu'au niveau de la fenêtre ronde. La mise en mouvement du canal cochléaire, et plus particulièrement de la membrane basilaire, provoque un fléchissement rythmique des cils des cellules sensorielles.
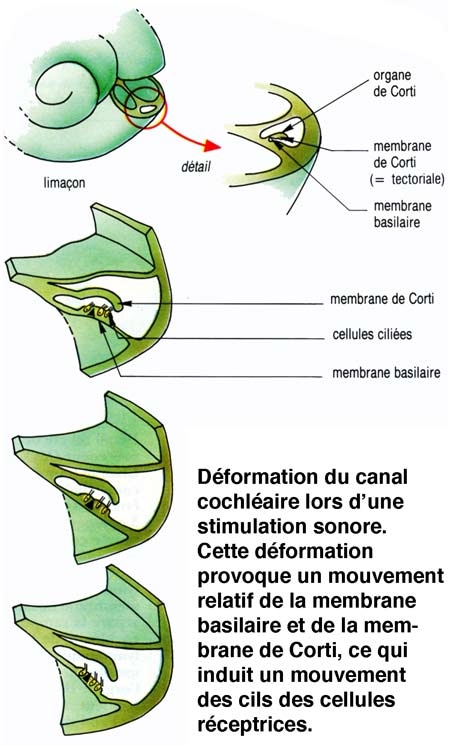
Le canal central contient l'organe sensoriel de l'audition, l'organe de Corti, tapissé des cellules sensorielles auditives : les cellules ciliées (15 000 par cochlée). La rampe vestibulaire et la rampe tympanique contiennent de la périlymphe riche en Na+, et encadrent le canal cochléaire qui, lui, contient de l'endolymphe riche en K+. Les cellules ciliées sont coiffées d'une touffe de stéréocils, qui transforment l'onde sonore en signal électrique. La partie basale de la membrane basilaire, peu large et épaisse, vibre préférentiellement pour des fréquences sonores élevées. À l'opposé, la région terminale (côté hélicotrème) vibre à des fréquences sonores faibles. Le codage de la fréquence sonore est donc directement dépendant de la position des cellules ciliées le long de la cochlée. La rampe tympanique et le canal cochléaire sont séparés par la membrane basilaire qui porte l'organe sensoriel proprement dit, ou organe de Corti. Ce dernier contient des cellules réceptrices ciliées, organisées en trois rangées externes et une rangée interne. L'ensemble est recouvert d'une membrane tectoriale attachée au côté interne de la cochlée.
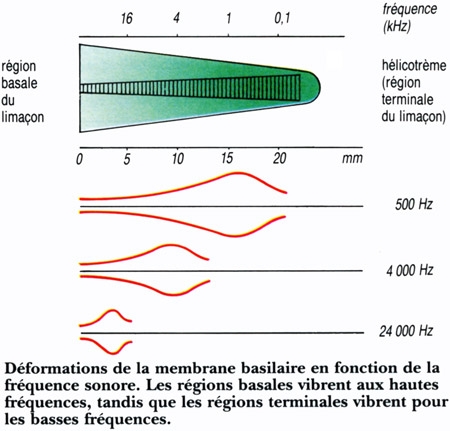
L'onde sonore amplifiée par l'oreille moyenne est en effet transformée en onde liquidienne qui va mettre en vibration la touffe ciliaire des cellules ciliées. Ces cellules sont des transducteurs : elles transforment les mouvements de leurs stéréocils en signal nerveux transmis au nerf auditif. Le long de la cochlée, chaque cellule ciliée répond préférentiellement à une fréquence sonore donnée, pour permettre au cerveau de différencier la hauteur des sons (batterie de résonateurs) : dans la partie basale de la cochlée sont codés les sons aigus, au sommet les sons graves.
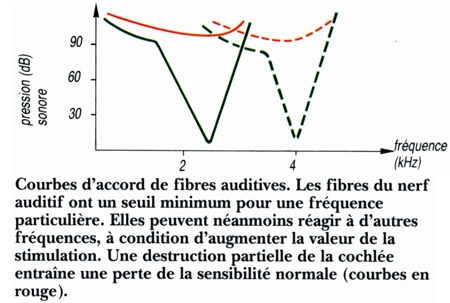
2. Les cellules ciliées assurent le codage de l'information
La différence de potentiel transmembranaire des cellules ciliées varie de façon synchrone avec le mouvement des cils. Au cours du mouvement des cils vers remplacement équivalent du kinocil, la membrane de leur extrémité se déforme, et permet l'ouverture de canaux K+. Les cils baignant dans une endolymphe riche en K+, un courant entrant d'ions positifs (K+) s'établit, provoquant une dépolarisation de la membrane. Cette dépolarisation provoque l'entrée d'ions Ca2+ qui permettent à la fois le mouvement des filaments contractiles de la plaque cuticulaire (ce qui accentue le mouvement des cils) et l'ouverture de canaux K+ situés sur la membrane du corps cellulaire. Ces derniers étant ouverts vers la périlymphe, le courant K+ qui s'établit est un courant sortant, repolarisant la membrane. Le potentiel générateur et les potentiels d'action apparaissent au niveau postsynaptique, dans la fibre sensorielle.
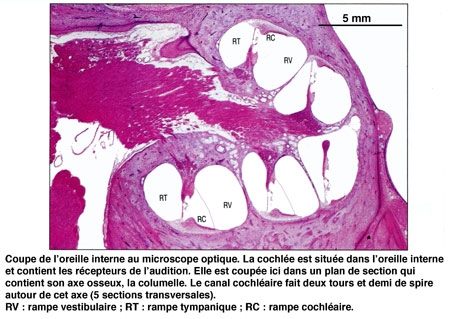
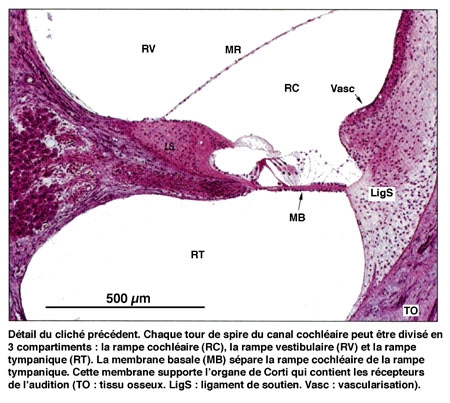
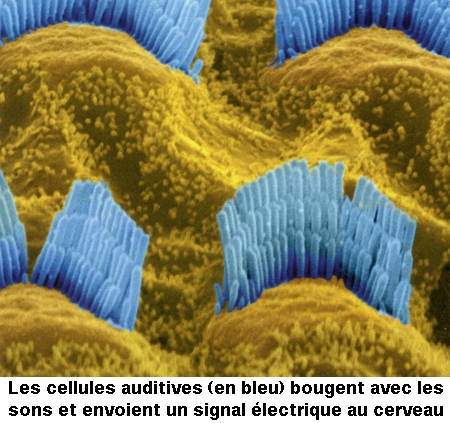
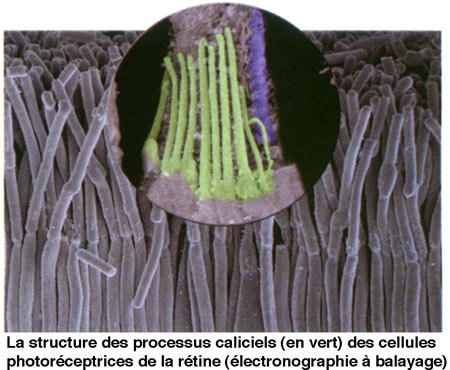
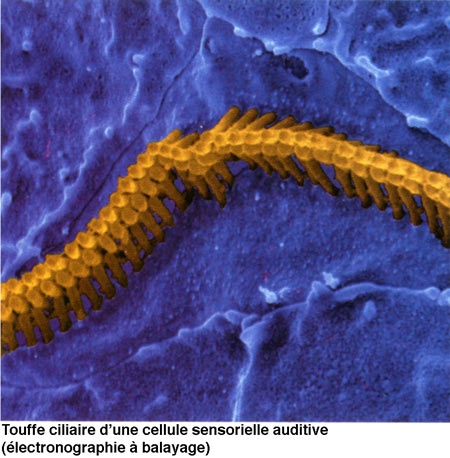
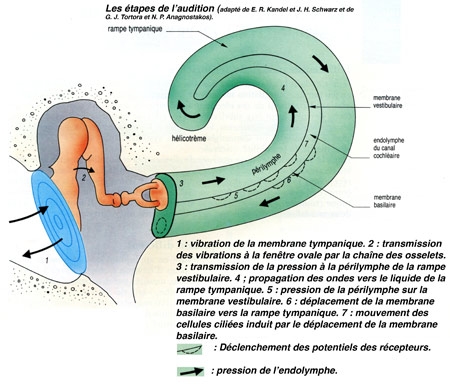
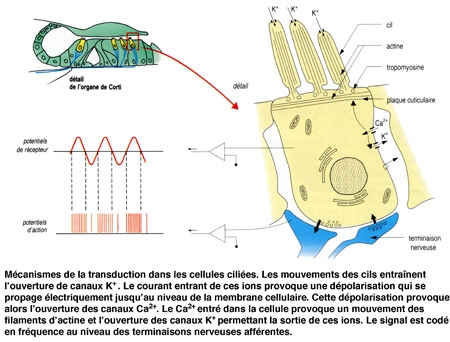
L'oreille interne contient, outre l'organe de l'ouïe, le vestibule, organe de perception de l'équilibre, repérant la position angulaire de la tête et les mouvements d'accélération.
3. Le cerveau traite les signaux
Le cerveau interprète le signal comme un son de la hauteur tonale correspondant au groupe de cellules excitées. L'analyse de l'intensité du son repose, quant à elle, sur divers mécanismes selon la fréquence sonore, le taux de décharges des neurones auditifs et la nature des neurones qui répondent (neurones à haut seuil ou à bas seuil).
Chaque fibre sensorielle est préférentiellement mise enjeu pour une fréquence particulière, mais peut également être mise en jeu par des fréquences différentes. La courbe du seuil de ces fibres en fonction de la fréquence sonore constitue leur courbe d'accord. Au niveau du cortex auditif les cellules sont organisées en colonnes fonctionnelles. Les neurones appartenant à une même colonne ont tous des courbes d'accord semblables. Les informations auditives liées au langage sont ensuite traitées dans des régions corticales particulières. Les aires principales impliquées dans ce traitement sont localisées normalement à l'hémisphère gauche. Ce sont l'aire temporale du langage, encore appelée aire de Wernicke, et l'aire frontale du langage, ou aire de Broca. La première est impliquée dans les processus sensoriels de reconnaissance de la parole, tandis que la seconde est plus directement liée à l'expression orale.
IV. Les surdités
- Origine des surdités
S'il existe quelques rares surdités d'origine centrale (impliquant le cerveau), la grande majorité des surdités est liée à un défaut de l'oreille elle-même.
Les surdités de transmission ont pour origine un défaut de l'oreille externe ou moyenne.
Les surdités neurosensorielles (surdités de perception) ont essentiellement pour origine une atteinte de la cochlée.
Dans ces deux catégories, certaines surdités sont génétiques, d'autres acquises.
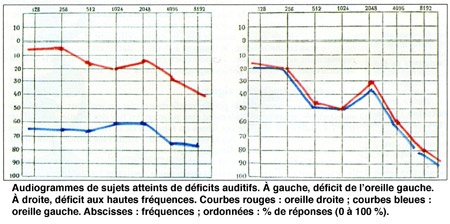
- Surdité : la part des gènes
L'audition est un outil majeur de communication, d'autant plus qu'elle est essentielle à l'acquisition de la parole : l'afflux d'informations auditives vers les «aires cérébrales de traitement du langage » est capital dans le jeune âge (au cours des six premières années de la vie - et tout particulièrement entre 0 et 3 ans) pour qu'un enfant apprenne à parler. Si la perception de la parole ou de la musique est une activité essentiellement cérébrale, la première étape du traitement des signaux sonores est effectuée par le système auditif périphérique. En cas de déficit non corrigé des atteintes auditives, le langage oral ne peut être acquis.
- Un millier d'enfants naît sourd chaque année et 65% des plus de 65 ans sont malentendants
Or en France, la surdité sévère ou profonde touche chaque année près d'un millier de nouveau-nés, avec les conséquences mentionnées sur l'acquisition du langage oral et sur le développement du lien du tout-petit avec ses proches. Un individu sur mille est de plus affecté par la surdité plus tard au cours de l'enfance. Ensuite, le pourcentage de malentendants dans la population ne cesse de progresser : 6% entre 15 et 24 ans, 9% entre 25 et 34 ans, 18% entre 35 et 44 ans et plus de 65% après 65 ans. Au total, la surdité concerne plusieurs millions de Français, à divers degrés de perte auditive, avec des conséquences variables sur leur vie sociale, Au-delà des surdités légères, les surdités moyennes ont, chez l'enfant, un impact négatif sur l'apprentissage scolaire, le développement cognitif et l'adaptation sociale. On l'a vu, les surdités profondes contrarient chez lui l'acquisition du langage oral et ont, chez l'adulte, l'isolement social pour conséquence majeure.
Face à ce problème considérable, la génétique a totalement modifié la donne, permettant une meilleure prise en charge des surdités de l'enfant, préparant pour lui de futures thérapies, et plus récemment, apportant un espoir supplémentaire : celui de prévenir les surdités liées au vieillissement.
- Surdités tardives : des problèmes de perception des fréquences aux acouphènes
« On peut imaginer l'organe sensoriel auditif comme un clavier de piano, où chaque région traite une fréquence qui lui est spécifique », explique Aziz El Amraoui, de l'unité Génétique et physiologie de l'audition, à l'Institut Pasteur. « La cochlée décompose les sons en ses fréquences élémentaires : les sons aigus (hautes fréquences) sont captées à la base de la spirale cochléaire. les sons graves (basses fréquences) dans sa partie haute. Avec le temps, les sons les plus aigus sont moins bien entendus ». La perte de l'audition avec l'âge s'accompagne d'une difficulté à distinguer les sons, à entendre par exemple une conversation dans un brouhaha. De plus, les surdités acquises sont assez fréquemment accompagnées d'acouphènes, des perceptions auditives (battements, grésillements, sifflements) en l'absence de stimulus externe, souvent très invalidantes : en France, près de 5 millions de personnes en souffriraient. Il semble que la perte auditive s'accompagne d'une augmentation de l'activité spontanée le long des voies auditives du cerveau aboutissant à ces perceptions « fantômes ». « Un rétablissement des entrées sensorielles à un niveau similaire à celui qui prévalait avant la perte auditive réduirait le gain central et supprimerait ainsi les acouphènes », avance Arnaud Norena, de l'Université Aix-Marseille, dans la revue Biofutur (novembre 2012). « Cette prédiction est en partie corroborée par la réduction/suppression des acouphènes produites par les aides auditives, les implants cochléaires et la stimulation acoustique ciblée sur les fréquences de la perte auditive. » Une raison de plus, donc, pour améliorer le traitement des surdités...
V. Les appareils auditifs existants
- Les aides auditives
Avec les aides auditives, le son est capté par un ou plusieurs microphones et le signal traité par un microprocesseur, amplifié, puis réémis via un écouteur dans le conduit auditif externe.
- Les aides en conduction osseuse stimulent directement l'oreille interne à travers les os du crâne ; un microphone capte les vibrations sonores, transmises à l'os temporal par un vibrateur placé derrière l'oreille ; les vibrations de la paroi osseuse autour de la cochlée sont ensuite transférées aux cellules sensorielles. Ces aides auditives sont parfois insuffisantes ou inadaptées à certaines formes de surdité. Deux sortes d'implants peuvent alors optimiser une audition résiduelle ou suppléer des structures défaillantes.
- L'implant d'oreille moyenne, fixé sur un osselet ou à proximité de l'oreille interne, capte les vibrations et les transmet à l'oreille interne.
- Avec l'implant cochléaire, un processeur externe transforme les sons en signaux électriques et les transmet par ondes à la partie interne implantée chirurgicalement, composée d'un ensemble d'électrodes. Ces dernières délivrent des impulsions électriques qui stimulent les fibres du nerf auditif.
- Les trois-quarts des surdités de l'enfant sont d'origine génétique
Dans les années 90, on ne connaissait pas la cause du déficit auditif des enfants qui naissaient sourds. Une équipe de généticiens à l'Institut Pasteur (voir Entretien) localisait alors les premiers gènes associés à des surdités sur des chromosomes humains. Ils firent cette découverte grâce à des collaborations avec plusieurs pays du bassin méditerranéen qui leur permirent de regrouper des données sur de nombreux individus appartenant à différentes familles, caractérisées par une forte consanguinité. Après ces premières grandes analyses génétiques, l'équipe contribua à l'identification d'une trentaine de gènes associés à des surdités, et d'autres dans le monde lui emboîtèrent le pas : 70 gènes impliqués dans des surdités héréditaires sont aujourd'hui connus. L'un d'eux, comme l'ont montre les chercheurs pasteuriens dès 1997, est à lui seul responsable de la moitié de ces surdités [le gène de la connexine 26]. Et l'on sait aujourd'hui que les trois-quarts des surdités de l'enfant sont d'origine génétique, le reste étant notamment dû a des traumatismes ou infections pendant la grossesse.
- Détecter le plus tôt possible les surdités néonatales, pour pouvoir appareiller
Cette aventure scientifique a révolutionné la prise en charge des surdités de l'enfant. Aujourd'hui, grâce aux travaux des chercheurs, des tests de diagnostic moléculaire des surdités les plus fréquentes sont utilisés en routine dans de nombreux pays du monde. Il est en effet important de détecter le plus tôt possible les surdités congénitales. Connaître le gène en cause permet de décider s'il faudra appareiller ou non l'enfant (dans certains cas, heureusement rares, l'appareillage ne résoudra rien] : plus l'enfant pourra entendre tôt, plus le retentissement de la surdité sur ses aptitudes linguistiques sera faible. Les chercheurs ont par exemple montré en 2006 que chez les enfants dont la surdité était due à un défaut du gène de l'otoferline, une protéine nécessaire à la transmission du signal par les cellules sensorielles auditives aux neurones auditifs, la pose précoce d'un implant cochléaire était recommandée alors que les caractéristiques de leur perte auditive conduisaient à s'interroger sur le bénéfice possible de l'implant. Parallèlement à ces avancées, ces recherches ont permis de comprendre de façon plus approfondie le fonctionnement de notre appareil auditif, et tout particulièrement celui de la cochlée, l'organe sensoriel de l'audition logé dans l'oreille interne.
- Un espoir majeur : la thérapie par les gènes
Aujourd'hui, beaucoup d'espoirs sont fondés sur la thérapie génique. Il s'agit là de restaurer la fonction auditive à l'aide du transfert du gène normal [«gène-médicament»], afin de compenser la fonction défectueuse de ce même gène chez le malentendant. Les méthodes envisagées sont diverses, combinant par exemple thérapie cellulaire et thérapie génique : certaines passent par l'introduction du « gène-médicament » dans des cellules souches ensuite différenciées en cellules de la cochlée, puis ré-introduites dans l'organe sensoriel. Les premiers résultats expérimentaux encouragent les scientifiques à poursuivre l'exploration de ces voies thérapeutiques nouvelles. Mais le domaine de l'enfance ne sera pas le seul bénéficiaire de la « révolution génétique » des recherches sur les surdités. Il s'agit désormais d'empêcher la dégradation de l'appareil auditif chez l'adulte. Les surdités de l'adulte, acquises au cours de la vie, sont liées à des traumatismes sonores, des infections (environ 20% des cas sont dus à des otites chroniques), des accidents liés à l'hyperpression (plongée) ou encore à une toxicité médicamenteuse. La plupart de ces facteurs contribuent à la destruction progressive des cellules sensorielles de la cochlée, indispensables à l'audition, et non renouvelables. Le vieillissement contribue par lui-même à une perte auditive qui apparaît le plus souvent à partir de 50-60 ans, la presbyacousie, aujourd'hui irréversible.
- De même que nous avons chacun un « capital soleil », nous aurions aussi un « capital son »
Là encore, la génétique joue un rôle : il existe une grande variabilité dans la sensibilité individuelle à l'atteinte auditive par la surexposition au bruit comme au traumatisme acoustique. Ainsi, chacun d'entre nous serait - selon son ADN - plus ou moins sensible au bruit. De même que nous avons chacun un « capital soleil », nous aurions aussi un « capital son ». Des gènes de prédisposition interviennent également dans notre sensibilité aux médicaments toxiques pour le système auditif (médicaments « ototoxiques »). Les scientifiques sont désormais engagés dans une nouvelle voie de recherche : comprendre comment notre oreille s'abîme, comment elle vieillit. La génétique est un outil majeur dans cette quête. Les réponses trouvées par les chercheurs devraient ouvrir la voie à des solutions thérapeutiques, que ce soit par traitements pharmacologique, génique ou cellulaire, associés ou non à des appareillages « classiques » du type implant cochléaire.
À ce jour - où le diagnostic génétique d'une surdité chez l'enfant est désormais une réalité - la perspective de prévenir l'apparition des surdités liées à l'âge n'a plus rien d'utopique...
- Face à un syndrome de surdité-cécité : l'espoir de traiter la cécité
L'équipe du Pr Christine Petit à l'Institut Pasteur étudie depuis quelques années une maladie génétique heureusement rare (elle touche une personne sur 10 000) qui associe surdité et cécité : le syndrome de Usher. Les enfants touchés naissent sourds et leur vision est progressivement altérée en raison du développement d'une rétinite pigmentaire. Parmi les différentes formes de la maladie, le type 1 (40% des cas) est le plus sévère : la surdité est profonde et les troubles de la vision apparaissent précocement. S'il existe une bonne prise en charge des patients concernant les troubles auditifs, - notamment grâce aux travaux des chercheurs et des équipes médicales -, aucun traitement ne permet aujourd'hui de stopper l'issue de la rétinite pigmentaire. Or les travaux du Pr Christine Petit[1], en collaboration avec le Pr José-Alain Sahel (Institut de la Vision), ont récemment permis de découvrir l'origine de la rétinite pigmentaire qui touche les patients atteints. Il s'agit d'un défaut dans l'organisation d'édifices cellulaires indispensables au maintien de la vision, les processus caliciels (voir photo) des cellules photoréceptrices de la rétine, dû au dysfonctionnement de l'une quelconque des cinq protéines impliquées dans cette maladie, et qui, ensemble, assurent la cohésion de ces structures. Ces travaux qui définissent les cibles de la maladie vont guider les approches thérapeutiques, aujourd'hui en cours, de cette cécité.
L'interview du Pr Christine Petit
Christine Petit est professeure au Collège de France et à l'Institut Pasteur, responsable de l'unité de Génétique et physiologie de l'audition.
Notre nouvel objectif est de comprendre les processus défectueux dans la surdité tardive pour en prévenir la survenue et la progression.
"Le domaine des surdités héréditaires de l'enfant a été transformé par les recherches de notre équipe."
Comment est née l'approche génétique de l'audition ? L'audition a été initialement explorée par les physiciens - intrigués par les performances exceptionnelles de l'organe sensoriel, la cochlée, qui se comporte comme un analyseur fréquentiel, transforme les sons, signaux mécaniques, en signaux électriques, les amplifie et aussi les distord... -, puis par les physiologistes, et enfin, par les généticiens. Nous avons mis en œuvre une approche génétique de l'audition pour déchiffrer les bases cellulaires et moléculaires du développement et du fonctionnement de la cochlée, qui échappaient jusque là à toute analyse en raison du tout petit nombre de cellules cochléaires. La cochlée ne comporte en effet que quelques milliers de cellules sensorielles auditives. La rétine, pour comparaison, est dotée d'une centaine de millions de cellules sensorielles photoréceptrices. L'intérêt de rapproche génétique pour identifier les molécules requises pour une fonction cochléaire puis en déchiffrer les mécanismes moléculaires correspondants tient au fait que son efficacité est indépendante du nombre de cellules ou de molécules impliquées dans la fonction explorée.
Qu'avez-vous découvert ?
En localisant les premiers gènes responsables de cette atteinte sensorielle sur les chromosomes humains, dans les années 1990, nous avons ouvert le domaine de l'audition à l'analyse génétique. Puis nous avons développé une approche spécifique pour identifier les gènes dont l'atteinte était susceptible d'être à l'origine de surdité chez l'enfant. Cette démarche nous a conduit à la découverte rapide de plusieurs gènes responsables de surdité précoce. Aujourd'hui, 70 de ces gènes sont connus. Par le développement d'approches pluridisciplinaires, impliquant à côté des généticiens, des biologistes cellulaires, des biochimistes, des électrophysiologistes et des physiciens, nous nous sommes ensuite engagés dans l'étude des mécanismes défectueux dans ces surdités. Nous avons pu en parallèle, déchiffrer les mécanismes moléculaires qui sous-tendent certaines propriétés physiologiques connues de la cochlée et éclairer les rôles de plusieurs structures cochléaires. Un grand nombre de laboratoires à travers le monde se sont ensuite engagés dans ce champ disciplinaire. C'est un domaine scientifique et médical particulièrement dynamique.
Quelles applications ces découvertes ont-elles permis pour les surdités de l'enfant ?
Le développement du diagnostic moléculaire de surdité héréditaire permet d'informer les familles qui le souhaitent sur le risque de récurrence d'une atteinte auditive chez les enfants à venir. La connaissance de la pathogénie des diverses formes de surdité permet, dans certains cas, de recommander l'utilisation d'appareils auditifs, en particulier de l'implantation cochléaire, alors que les caractéristiques audiologiques de la surdité suscitent le doute sur leur efficacité. Elle permet parfois, à l'inverse, de conseiller d'éviter tout appareillage auditif et de s'en remettre à des traitements pharmacologiques, dont la connaissance des mécanismes pathogéniques de ces surdités permet de prédire l'efficacité. Aujourd'hui, fondés sur ces avancées, des espoirs de traitements se profilent. Plusieurs laboratoires à travers le monde sont engagés dans la recherche de traitements des surdités, en particulier par voie génétique.
Vous vous intéressez aussi aux surdités tardives. Que peut-on espérer ?
Notre objectif est de comprendre les processus défectueux dans la surdité tardive pour en prévenir la survenue et la progression. On sait que certains gènes prédisposent à la presbyacousie, et aux atteintes auditives liées à une surexposition au bruit. La plupart d'entre eux restent à découvrir. C'est une étape incontournable pour caractériser les processus pathogéniques en cause. Nous identifions par exemple un nombre croissant d'atteintes auditives liées à des perturbations du métabolisme antioxydant que l'on pourrait vraisemblablement prévenir par des médicaments existants. Dans ce but, nous avons créé au cours des dernières années un réseau national qui comporte plusieurs services d'ORL afin de regrouper des familles atteintes de presbyacousie pour rechercher les gènes impliqués. Une analyse approfondie de leur audition a été mise en place. Elle comporte des tests psychoacoustiques pour nous permettre de reconnaître les atteintes de l'intelligibilité de la parole qui sont souvent, chez ces personnes, beaucoup plus sérieuses que ne le prédit la mesure du seuil de leur perception auditive par l'audiogramme. C'est un effort collectif important. Il nous permet aujourd'hui d'analyser environ 150 familles, auxquelles s'ajoute plus d'un millier d'individus atteints. On imagine aisément que le coût de cette recherche, qui repose sur l'analyse exhaustive des génomes de ces personnes, est très élevé.
Glossaire :
Courbe isophone : courbe reliant, sur un audiogramme, les valeurs de pressions sonores entraînant des sensations d'intensité identique.
Décibel (dB) : n.m., unité de pression sonore relative. 1 dB = 20 log.(P/2.10-5 ), où P est la pression sonore exprimée en newton.m-2.
Phone : n. m., unité sans dimension permettant de graduer l'échelle de sensibilité auditive en fonction de la fréquence des sons. La référence se situe à 1 kHz où 1 phone = 1dB.
Cochlée : n.f., partie de l'oreille interne formée de l'enroulement de trois canaux et dans laquelle sont situées les cellules réceptrices sensibles aux vibrations sonores.
Hélicotrème : n.m., extrémité apicale de la cochlée.
Organe de Corti : partie de la cochlée regroupant les cellules sensorielles.
Tympan : n.m., membrane séparant l'oreille externe de l'oreille moyenne.
[1] Menés avec les Drs Azizi El Amraoui et Iman Sahly.
Sources :
- Surdités La lettre de l'Institut Pasteur, revue trimestrielle février 2013.
- Périlleux E., Anselme B., Richard D. (1995). - Biologie humaine, anatomie, physiologie, santé. Ed. Nathan.
18:20 Publié dans Biologie, Environnement-Écologie, Médecine | Tags : audition, oreille, surdité, institut pasteur | Lien permanent | Commentaires (0) | Facebook | | |
18/01/2013
Le ver, la mouche et la souris : trois animaux d'intérêt pour la biologie du développement
Le ver, la mouche et la souris : trois animaux d'intérêt pour la biologie du développement
par
Claude-Roland Marchand
(professeur honoraire de l'Université de Franche-Comté)
La compréhension du vivant ne peut se faire qu'en observant la diversité des espèces ; leurs affinités phénotypiques et reproductrices permettant de les regrouper ; leur physiologie et leur dynamique vitale les rapprochant ou les singularisant. Au cours des siècles, l'intérêt s'est porté sur des animaux de proximité, sauvages ou domestiques, dans divers milieux, et les conclusions qu'on en tirait étaient nourries d'une logique et d'une culture très subjectives, imprégnées de croyances philosophiques ou religieuses (L'Homme faisait l'objet d'une lointaine et prudente approche, car il n'était pas considéré comme un animal !).
Le XXe siècle a vu l'essor de l'embryologie expérimentale qui s'est intéressée au développement animal, depuis la cellule-œuf jusqu'à l'animal différencié et fonctionnel, en utilisant des Invertébrés (Hydre, Planaire, Oursin...) ou des Vertébrés (Grenouille, Poulet, Souris).
Quels sont les intérêts des modèles animaux ?
Les Unicellulaires utilisés en microbiologie nous ont renseignés sur les propriétés et les fonctions de la cellule. Mais le vivant comporte également des animaux complexes, les Métazoaires, étudiés depuis longtemps et de diverses manières. Ils demandent une approche particulière qui a évolué au cours des siècles en fonction des avancées techniques. On les a observés in situ mais également et différemment en laboratoire. Pour mémoire citons les plus célèbres : les Pinsons de Darwin, le Chien de Pavlov ou les Grenouilles de Rostand qui ont modifié notre vision de l'évolution, de la physiologie ou de la tératologie.
Nous évoquerons trois des principaux animaux-modèles utilisés au XXe siècle et encore aujourd'hui : un Ver Nématode Caenorhabditis elegans, un Insecte, la mouche du vinaigre, Drosophila melanogaster et un Mammifère, la souris, Mus musculus.
Nous verrons en quoi leur étude a fait progresser nos connaissances fondamentales et nous évoquerons les applications possibles qui pourraient être envisagées dans de nombreux domaines.
Le ver Nématode CAENORHABDITIS elegans
Les « pères » de ce modèle, utilisé depuis les années 60, Sydney Brenner, John Sulston et Bob Horvitz ont reçu le prix Nobel en 2002.
C'est un Nématode (Ascarides) de 1 mm de longueur. 20 jours de vie, une génération tous les 4 jours. Ce sont habituellement des hermaphrodites autoféconds qui contiennent 959 cellules et 1 fois sur 1000 des mâles formés de 1031 cellules. Les mâles peuvent s'accoupler avec les hermaphrodites, ce qui permet d'obtenir des lignées pures. Le séquençage de son génome, réalisé en 1998 (premier animal à être totalement séquencé) montre qu'il contient 19 099 gènes dont environ 200 peuvent être rendus létaux par mutation.
Le Nématode Caenorhabditis elegans
550 millions d'années séparent l'apparition de ce Ver et celle des Mammifères et pourtant il existe de nombreuses similitudes fonctionnelles au niveau neuronal ou au cours du vieillissement. Ces propriétés accentuent l'intérêt de l'étude de cet animal qui se nourrit d'Escherichia coli, est transparent, facile à observer sous le microscope et aisé à manipuler : ablation de cellules par laser, injection de marqueurs, de séquences d'ADN ou d'ARN antisens pour clarifier la fonction de tel ou tel gène à tel moment du développement.
On a pu établir chez lui le lignage complet de toutes les cellules issues de la fécondation. Sur la carte ainsi dressée, on peut retrouver l'origine de toutes les cellules constitutives de tel ou tel organe et le moment où la lignée germinale est isolée de la lignée somatique. C'était un rêve pour les embryologistes : il s'est réalisé à la fin du siècle dernier !
Outre le criblage génique, on a noté chez Caenorhabditis qu'une cellule sur 8 mourait au cours du développement normal. C'est une mort cellulaire programmée appelée apoptose qui se réalise grâce à l'action de protéines appelées caspases. Elle est contrôlée par les gènes ced3, ced4 qui, lorsqu'ils sont mutés, provoquent un excès de survie cellulaire. L'apoptose est une étape incontournable dans les processus d'organogenèse du vivant ; on en verra un exemple, plus loin, avec le développement des doigts chez les tétrapodes. À l'inverse le gène ced9 favorise la survie des cellules.
L'établissement du plan corporel, la symétrisation et l'organogenèse sont commandés par un jeu complexe et subtil de gènes qui s'expriment dans le temps et dans l'espace en étroite synergie. On a en particulier isolé les gènes egl5 et CePhox2/ceh17 qui participent à rétablissement de l'axe antéro-postérieur du ver, et de la mise en place des cellules nerveuses.
En outre, le séquençage de son génome révèle la présence de gènes homologues de psi, park2 et insr responsables respectivement des maladies d'Alzheimer, de Parkinson et du diabète de type 2 chez l'Homme.
À titre expérimental, 3 millions de Caenorhabditis ont été placés dans une navette spatiale Columbia en 2003 afin d'observer le déroulement des différentes étapes du développement en apesanteur. Après l'accident de cette navette, on a retrouvé des vers vivants parmi les débris, dans leurs boîtes d'élevage, révélant de manière inattendue la relative résistance de ces vers.
Deuxième animal-modèle : La mouche des fruits, DROSOPHILA melanogaster
Tout a commencé, pour cette Mouche aux yeux rouges, avec le naturaliste américain Thomas Morgan (1866-1945), prix Nobel 1933, qui a voulu vérifier avec elle, les lois de Mendel redécouvertes au début du XXe siècle. Elle est très fertile, facile à élever, facile à observer et présente souvent des mutations. Son génome comporte 13 000 gènes dont plus de 75% sont homologues aux gènes humains.

La Mouche des fruits : Drosophila melanogaster
Dès 1927 C. Bridges observe des Drosophiles mutantes présentant quatre paires de pattes, ou deux paires d'ailes. En 1954, E. Lewis observera d'autres mutations. Ces deux chercheurs leur donnent des noms évocateurs : bithoraxoïd, bithorax, postbithorax, ultrabithorax. Et lorsque des antennes sont transformées en pattes Lewis parle d'Antennapedia. Dès lors la notion d'homéose est proposée : elle désigne, selon Bateson, en 1894, la transformation d'un organe en un autre. Lewis et ses collaborateurs parleront désormais de gènes homéotiques, ou homéogènes à l'origine de ces transformations spectaculaires.
Lewis localisera ces homéogènes sur le chromosome 3 de la Drosophile. Chaque homéogène comporte une séquence de 180 paires de bases, l'homéobox, qu'on retrouve également chez le Xénope, la Souris et chez l'Homme.
De 1983 à 1990, E. Lewis et ses collaborateurs, Christiane Nüsslein-Volhard et Eric Wieschaus, affineront leurs découvertes. Ce qui leur valut le Prix Nobel en 1995. Cette découverte montre que des gènes majeurs, conservés au cours de révolution, contrôlent le schéma corporel de base de chaque zootype ; c'est un paradigme qui s'avère valable pour le règne végétal.
Quelques images nous permettent de suivre les différentes étapes du développement de la Drosophile et de localiser les gènes architectes qui contrôlent l'organogenèse, dans l'espace et dans le temps.
1 = Les polarités dorso-ventrale et antéro-postérieure
sont esquissées dans les ovarioles de la femelle et confirmées après la fécondation. Un grand nombre de gènes (dont nous ne donnerons pas la longue liste) participent à rétablissement de l'identité des territoires, comme par exemple dorsal, nanos, toll, torpedo, cactus… dont l'activité produit une combinaison de facteurs de transcription morphogénétiques. Sur ces schémas, on peut noter la répartition, selon un gradient, des produits des gènes bicoid, torso et nanos. Leur mutation conduit à des larves transformées : sans abdomen, sans tête, sans telson... Grâce aux anomalies, on peut localiser le moment et la région où s'expriment les gènes d'intérêts.
2 = L'identité segmentaire se réalise progressivement, en s'affinant peu à peu, pour aboutir à une zonation conforme au plan de l'Insecte : tête, thorax, abdomen. On ne peut manquer d'être surpris par ce qui peut paraître comme une certaine logique dans le déroulement de ces étapes : du simple au complexe et de l'avant vers l'arrière.
Trois groupes de gènes, les gènes gap[1], pair-rule[2], de polarité segmentaire interagissent et construisent le plan corporel segmentaire.
La mise en évidence de leurs interrelations et de leurs domaines d'expression, parfois chevauchants, est le résultat d'un travail minutieux de biologie moléculaire, intéressant pour la Drosophile, mais aussi au plan fondamental.
3 = Les homéogènes : Attardons-nous un peu sur ce que l'étude approfondie des mutations de la Drosophile a apporté à la connaissance des gènes architectes.
Les plus spectaculaires sont des mouches porteuses de 4 ailes (mutation appelée Ultrabithorax) et des mouches dont les antennes sont transformées en pattes (mutation appelée Antennapedia).
Le mérite de Lewis a été de localiser, en 1978, les gènes responsables de ces mutations. Ils se trouvent tous sur le chromosome 3. En 1983 il parvient avec son équipe à isoler et séquencer ces gènes qu'il range désormais dans le groupe des gènes homéotiques. Car, nous le répétons, leur expression ectopique construit un organe conforme, mais qui n'est pas à sa place : des ailes à la place des balanciers ou une patte à la place de l'antenne.
Voici le schéma qu'il propose, sur le même chromosome 3 (colinéarité), de l'avant à l'arrière 8 homéogènes regroupés en deux ensembles :
5 dans l'ensemble ANT : lab, pb, Dfd, Scr, Antp.
3 dans l'ensemble BX : Ubx, abd-A, Abd-B.
Les abréviations correspondent à la caractéristique d'une mutation du gène concerné.
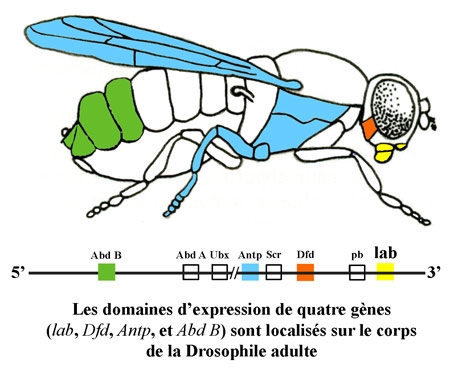
(Abréviations : lab = labial ; pb = proboscipedia ; Dfd = deformed ; Scr = sex comb reduced ; Antp = antennapedia ; Ubx = ultrabithorax ; Abd A = abdominal A ; Abd B = abdominal B).
Si on délimite sur le plan du corps de l'animal les domaines d'expression des principaux gènes identifiés, on s'aperçoit que l'identité segmentaire est souvent, sauf pour l'extrémité antérieure, le résultat d'une action conjuguée de plusieurs produits géniques. Cette synergie notée chez la Drosophile existe également chez les autres métazoaires, y compris les Mammifères comme on le verra plus loin.
Voyons maintenant les mutations concernant les yeux de la Drosophile.
En 1915, M. Hoge observait déjà des mouches sans yeux ou avec des yeux réduits. Cette anomalie provenait d'un gène défaillant appelé Eyeless isolé en 1993, deux ans après qu'un homologue appelé Small Eye / Aniridia eût été identifié chez la Souris. Bien que l'œil simple de Mammifères ait peu de ressemblances morphologiques avec l'œil composé d'un Insecte, il semblerait que sa construction utilise des procédés similaires, avec probablement la mise en jeu de plus de 2000 gènes ! C'est en 1995 que W. J. Gehring et son équipe parviennent à activer Eyeless et à obtenir une mouche avec des yeux sur les pattes et sur les antennes.
Finalement ces différents gènes ont été placés dans le groupe PAX6, en raison de leur structure. Nous n'omettrons pas de mentionner la réussite d'une expérience inattendue : si on introduit le gène PAX6 de la Souris dans une larve de Drosophile on induit la formation d'un œil supplémentaire de type Insecte. Ce qui signifie que le code du gène architecte de la Souris est « compris » par la Mouche et est ensuite « traduit » en organe de mouche. Voici donc levé un tabou qui posait problème à Darwin : il n'osait pas imaginer la construction de l'œil à partir d'un même prototype soumis ensuite à la sélection naturelle. La biologie moléculaire démontre donc l'existence de mécanismes identiques, malgré la distance phylogénétique ; ceci est d'autant plus vrai que l'équipe de Gehring a trouvé des homologues de Eyeless et de Small eye chez un Ver Némerte, une Planaire et le Calmar !
Après ce survol des résultats obtenus en biologie moléculaire expérimentale, nous pressentons qu'au-delà du Ver ou de la Mouche nous allons vers une universalité de certains processus, de certains signaux qui s'accompagnent d'une conservation de leurs patrons d'expression et des mécanismes qui les initient et les régulent.
Voyons avec le troisième animal de laboratoire si nos concepts seront confirmés ou infirmés.
La souris : MUS musculus
Voici le Mammifère le plus facile à élever, véritable « cobaye » de la recherche mondiale : de petite taille, un cycle toutes les six semaines, une gestation courte de 21 jours, 6 à 12 petits par portée. La sélection a permis d'obtenir d'elle des lignées pures qui permettent de réaliser de manière reproductible, des manipulations génétiques comme des transferts, des suppressions, des amplifications de gènes ou une modélisation de maladies humaines. La transgénèse est effectuée sur des œufs prélevés juste après la fécondation. L'ADN injecté peut être inséré dans tous les noyaux et sera exprimé dans la descendance.

La Souris : Mus musculus
La biologie moléculaire a séquencé la totalité des 40 chromosomes de la Souris en 2003 ; il en ressort que 98 % de ses gènes sont homologues à ceux de l'Homme, ce qui constitue un matériel propice à l'étude des maladies génétiques, de certains cancers, mais aussi de la régulation des étapes-clés de l'embryogénèse. Parmi les découvertes spectaculaires dues à cette technique mentionnons le repérage, le séquençage et la localisation de 41 homéogènes répartis sur 4 chromosomes et présentant de nombreuses homologies avec les 8 homéogènes de la Drosophile.
Sur le schéma suivant on peut voir leur position et quelques-uns de leurs domaines d'expression. Ces homéogènes sont désignés par les termes Hox a, Hox b, Hox c, Hox d, suivis d'un numéro qui fait référence à leurs positions respectives sur les chromosomes 6, 11, 15, 4.
Hox a1, Hox b1 et Hox d1 sont des homologues de lab de la Drosophile.
Hox a9, Hox b9, Hox c9 et Hox d9 sont homologues de abd B.
Ils commandent, en synergie, en interactivité, l'identité de certaines portions de l'embryon comme le montre le schéma simplifié d'un jeune fœtus.
Hox a13, Hox b13, Hox c13, Hox d13 sont des gènes paralogues. Ils peuvent agir de manière coordonnée, ou isolément, laissant supposer une certaine redondance, sorte de mémoire-secours.
Au plan évolutif on pense que ces gènes proviennent de plusieurs duplications à partir d'un gène ancestral dont ils ont conservé des séquences pendant des millions d'années.
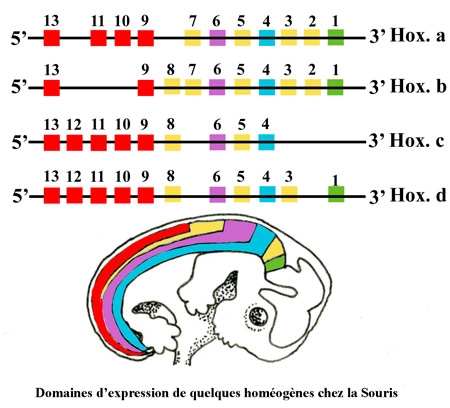
Mêmes figurés que ceux utilisés dans le schéma de la Drosophile présenté plus haut permettant d'établir les homologies. (N.B. : La transcription se fait toujours de 3' vers 5').
On pourrait présenter la même répartition des homéogènes localisés sur 4 des 46 chromosomes humains :
11 sur le chromosome 7 = HOX A
10 sur le chromosome 17 = HOX B
9 sur le chromosome 12 = HOX C
11 sur le chromosome 2 = HOX D
Ces gènes sont activés à un moment précis du développement embryonnaire par des signaux complexes et multiples ; comme chez la Drosophile les plus antérieurs s'expriment les premiers et s'appliquent à une zone antérieure ; les suivants leur succèdent, chevauchant ou non. Et cette combinatoire a pour résultat un segment, une zone, une ébauche qui se différenciera progressivement par la suite. On notera que les frontières antérieures d'expression sont toujours très nettes et qu'elles s'estompent vers l'arrière (sur la figure projetée on a l'exemple de Hox b1, Hox b4, Hox b5). Logiquement la redondance n'est pas exclue.
Exemple de l'implication des homéogènes dans la morphogénèse du membre : À partir de quatre bourgeons s'édifient les membres de la Souris dont les orientations antéro-postérieure, dorso-ventrale et proximo-distale s'établissent grâce à l'action combinée de plusieurs gènes, qui imposent également la mise en place des ébauches osseuses. La formation des espaces interdigitaux est réalisée grâce à la mort cellulaire programmée (apoptose) qui s'exerce sur des territoires précis de la palette distale.
Examinant les domaines d'expression de Hox d1 à Hox d13 à la fois sur le corps entier du fœtus et sur les bourgeons des pattes, on confirme la hiérarchie de la répartition des gènes et une certaine synergie entre eux. (L'extrémité du membre résulte d'une addition des produits d'au moins cinq gènes).
La mise en évidence de cette combinatoire a constitué un grand pas pour la biologie du développement ; elle a permis d'expliquer l'origine de plusieurs anomalies (amélie, phocomélie... adactylie, acheirie...) et de prendre des mesures de prévention.
Ces avancées moléculaires ouvrent des perspectives intéressantes, dans la mesure où ce qui est possible chez la Souris pourrait bien être, à terme et avec toutes les précautions qui s'imposent, tenté chez l'Homme.
Citons quelques exemples de modifications obtenues chez la Souris par transgénèse : inhiber l'expression d'un gène à la base d'un bourgeon de membre, ajouter des vertèbres cervicales, transformer une vertèbre lombaire en vertèbre thoracique, ajouter une paire de côtes... Dernières images de cette capacité à modifier le vivant : si on empêche l'expression de PAX 6 chez la souris : pas d'œil ou œil atypique.
Si on place Hox d9 dans la mouche : l'antenne est transformée en patte (d'insecte).
*
* * *
Ces résultats illustrent les possibilités de la biologie moléculaire et peuvent éveiller de réels espoirs pour la thérapie génique régulièrement évoquée lorsque des résultats prometteurs sont obtenus sur des souris transgéniques exprimant des maladies humaines comme certains cancers, la maladie d'Alzheimer, de Parkinson, le diabète ou les myopathies.
*
* * *
La lignée de souris Tg2576 constitue un modèle de référence pour la maladie d'Alzheimer, alors que les souris NOD (Non Obèse Diabetic) ou knock out Glut4 le sont pour le diabète.
Nous n'avons pas pu aborder les innombrables recherches effectuées sur les cellules-souches et sur le clonage dont les retombées ouvrent des perspectives prometteuses pour la santé humaine. Cela mériterait un exposé particulier, long et détaillé.
Et si l'on prolonge l'exploration moléculaire chez d'autres Vertébrés on peut comparer chez le Poulet et chez la Couleuvre les domaines d'expression d'Hox c6 (homéogène de Souris !).
Les frontières entre espèces semblent s'estomper et ceci nous amène à conclure.
CONCLUSIONS
Sans l'étude de ces trois animaux, nous n'aurions pas pu accéder à plusieurs processus cellulaires fondamentaux comme l'apoptose ; nous n'aurions pas pu connaître la combinatoire génique qui commande la morphogénèse embryonnaire.
Nous disposons donc de nouveaux concepts applicables à la recherche fondamentale et à la recherche appliquée, compte tenu de l'universalité du fonctionnement cellulaire, et des similitudes des processus de l'organogenèse.
Les homologies structurales et fonctionnelles nous permettent de vérifier que l'ontogenèse rappelle la phylogenèse, et que le génome des espèces a conservé une part de l'héritage d'un ancêtre commun. Du temps de Darwin, il était inconvenant d'imaginer que l'Homme descende du Singe... Qu'en serait-il aujourd'hui si on évoquait une lointaine filiation Homme/Insecte ?
L'étude d'animaux « modèles » nous paraît indispensable à la connaissance du fonctionnement cellulaire et organique, à la mise au point de thérapies innovantes et efficaces, sans oublier le positionnement de chaque espèce dans la chaîne évolutive.
Professeur Claude-Roland MARCHAND
(Communication présentée à l'Académie des Sciences et Belles Lettres de Besançon Franche-Comté le 6 avril 2009 )
09:40 Publié dans Biologie, Médecine | Lien permanent | Commentaires (1) | Facebook | | |
04/11/2012
Les OGM sont-ils des poisons ?
 Les OGM sont-ils des poisons ?
Les OGM sont-ils des poisons ?
(dernière mise à jour : 29/01/2013)
Des chercheurs français ont étudié secrètement, pendant deux ans, 200 rats nourris au maïs transgénique. Tumeurs, pathologies lourdes… une hécatombe. Et une bombe pour l'industrie OGM.
(Cet article signé Guillaume Malaurie est paru dans le "Nouvel Observateur" daté du 20 septembre 2012).
C’est une véritable bombe que lance, ce 19 septembre 2012 à 15 heures, la très sérieuse revue américaine "Food and Chemical Toxicology" - une référence en matière de toxicologie alimentaire - en publiant les résultats de l’expérimentation menée par l’équipe du français Gilles-Eric Séralini, professeur de biologie moléculaire à l'université de Caen. Une bombe à fragmentation : scientifique, sanitaire, politique et industrielle. Elle pulvérise en effet une vérité officielle : l’innocuité du maïs génétiquement modifié.
Lourdement toxique et souvent mortel
Même à faible dose, l’OGM étudié se révèle lourdement toxique et souvent mortel pour des rats. À tel point que, s’il s’agissait d’un médicament, il devrait être suspendu séance tenante dans l’attente de nouvelles investigations. Car c’est ce même OGM que l’on retrouve dans nos assiettes, à travers la viande, les œufs ou le lait.
En 2006, c’est comme un véritable thriller que commence cette recherche, dont le maître d’œuvre, Gilles-Eric Séralini, divulgue lui-même les conclusions dans un ouvrage à paraître la semaine prochaine ("Tous cobayes !", Flammarion, en librairie le 26 septembre 2012).
Nom de code In Vivo
Jusqu’en 2011, les chercheurs ont travaillé dans des conditions de quasi-clandestinité. Ils ont crypté leurs courriels comme au Pentagone, se sont interdit toute discussion téléphonique et ont même lancé une étude leurre tant ils craignaient un coup de Jarnac des multinationales de la semence.
Le récit de l’opération – nom de code In Vivo - évoque la très difficile récupération de semences de maïs OGM NK 603, propriété brevetée de Monsanto, par le truchement d’un lycée agricole canadien. Puis la récolte et le rapatriement des "gros sacs de jute" sur le port du Havre fin 2007, avant la fabrication de croquettes dans le secret le plus total et la sélection de deux cents rats de laboratoires dits Sprague Dawley. Bilan ? Glaçant : "Après moins d’un an de menus différenciés au maïs OGM, confie le professeur Séralini, c’était une hécatombe parmi nos rats, dont je n’avais pas imaginé l’ampleur".
Pathologies lourdes, tumeurs mammaires
Tous les groupes de rats, qu’ils soient nourris avec le maïs OGM traité ou non au Roundup, l'herbicide de Monsanto, ou encore alimentés avec une eau contenant de faibles doses d’herbicide présent dans les champs OGM, sont frappés par une multitude de pathologies lourdes au 13e mois de l’expérience. Chez les femelles, cela se manifeste par des explosions en chaine de tumeurs mammaires qui atteignent parfois jusqu’à 25% de leur poids. Chez les mâles, ce sont les organes dépurateurs, le foie et les reins, qui sont atteints d’anomalies marquées ou sévères. Avec une fréquence deux à cinq fois plus importante que pour les rongeurs nourris au maïs sans OGM.
Les rats nourris pendant deux avec du maïs OGM
ont développé de spectaculaires tumeurs
© J+B SÉQUENCES
Comparaison implacable : les rats OGM déclenchent donc de deux à trois fois plus de tumeurs que les rats non OGM quel que soit leur sexe. Au début du 24° mois, c’est-à-dire à la fin de leur vie, de 50% à 80% des femelles OGM sont touchées contre seulement 30% chez les non-OGM.
Surtout, les tumeurs surviennent nettement plus vite chez les rats OGM : vingt mois plus tôt chez les mâles, trois mois plus tôt chez les femelles. Pour un animal qui bénéficie de deux ans d’espérance de vie, l’écart est considérable. A titre de comparaison, un an pour un rongeur, c’est à peu près l’équivalent d’une quarantaine d’années pour un homme…
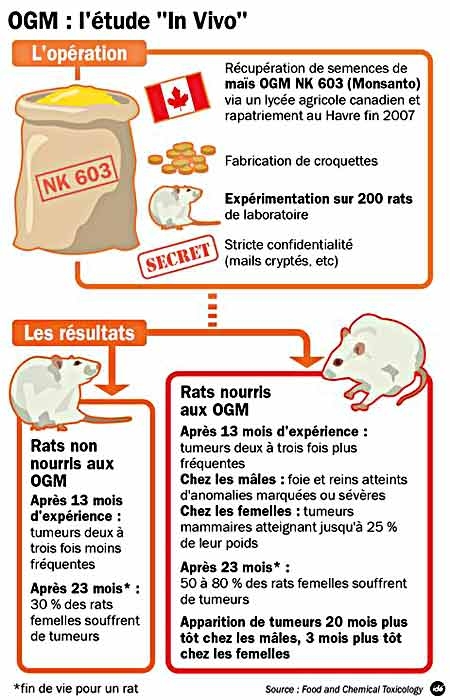
Exiger des comptes
C’est forte de ces conclusions que Corinne Lepage, dans un livre qui paraît vendredi 21 septembre ("La vérité sur les OGM, c’est notre affaire", Editions Charles Léopold Mayer), entend bien exiger des comptes auprès des politiques et des experts, français et européens, des agences sanitaires et de la Commission de Bruxelles, qui se sont si longtemps opposés et par tous les moyens au principe d’une étude de longue durée sur l'impact physiologique des OGM.
Cette bataille, l’ex-ministre de l’Écologie et première vice-présidente de la commission Environnement, Santé publique et Sécurité alimentaire à Strasbourg, la mène depuis quinze ans au sein du Criigen (Comité de Recherche et d’Information indépendantes sur le Génie génétique) avec Joël Spiroux et Gilles-Éric Séralini. Une simple association 1901 qui a pourtant été capable de réunir de bout en bout les fonds de cette recherche (3,2 millions d’euros) que ni l’Inra, ni le CNRS, ni aucun organisme public n’avaient jugé judicieux d’entreprendre.
Une étude financée par Auchan et Carrefour
Comment ? Autre surprise : en sollicitant la Fondation suisse Charles Léopold Mayer. Mais aussi les patrons de la grande distribution (Carrefour, Auchan..), qui se sont réunis pour l'occasion en association. Depuis la vache folle, ces derniers veulent en effet se prémunir de tout nouveau scandale alimentaire. À tel point que c’est Gérard Mulliez, fondateur du groupe Auchan, qui a débloqué les premiers financements.
L’étude du professeur Séralini laisse donc présager une nouvelle guerre meurtrière entre pro et anti-OGM. Les agences sanitaires exigeront-elles de toute urgence des études analogues pour vérifier les conclusions des scientifiques français ? Ce serait bien le moins. Monsanto, la plus grande firme mondiale de semences transgéniques, laissera-t-elle faire ? Peu probable : sa survie serait en jeu. Pour une seule plante OGM, il y a une centaine de variétés. Ce qui impliquerait au moins une dizaine d'études de 100 à 150 millions d’euros chacune !
Le temps de la vérité
Sauf que, dans cette nouvelle confrontation, le débat ne pourra plus s’enliser comme par le passé. Dés le 26 septembre, chacun pourra voir au cinéma le film choc de Jean-Paul Jaud, "Tous Cobayes ?", adapté du livre de Gilles-Éric Séralini, et les terribles images des rats étouffant dans leurs tumeurs. Des images qui vont faire le tour de la planète et d’internet, puisqu'elles ont été diffusées sur Canal+ (au "Grand Journal" du 19 septembre 2012) et le seront encore sur France 5 (le 16 octobre 2012 dans un documentaire). Pour les OGM, l’ère du doute s’achève. Le temps de la vérité commence.
À lire : l'interview de Gilles-Éric Séralini, ainsi que des extraits de son livre "Tous cobayes !" (Flammarion), dans "Le Nouvel Observateur" du 20 septembre 2012.
--
Roxanne MITRALIAS
Pôle Agriculture et environnement :
OGM, sanitaire, semences, biodiversité
Confédération Paysanne
104 rue Robespierre - 93170 Bagnolet
01.43.62.18.73
http://www.confederationpaysanne.fr
La démarche expérimentale de Gilles-Éric Séralini a été vertement critiquée. Voir également l'article résumant les remarques de différents organismes sur la méthode.
Un article signé Hervé Ratel dans Sciences et Avenir de novembre 2012, p. 40 expose les arguments de l'autorité Européenne de Sécurité des Aliments (Efsa) :
OGM : les points clés de l'étude controversée
Après la publication de l'étude du Pr Séralini sur les effets toxiques des OGM, analyse des éléments fondamentaux pour comprendre les questions en jeu.
Des tumeurs grosses comme des balles de ping-pong, des atteintes hépatiques et rénales graves, des longévités amputées... Photos chocs à l'appui, les révélations clé l'étude de toxicologie sur des rats nourris avec un maïs OGM, publiées dans le Nouvel Observateur du 19 septembre 2012, ont fait l'effet d'une bombe, et la une des journaux. Publié dans la revue Food and Chemical Toxicology, le travail dirigé par Gilles-Eric Séralini (université de Caen) est le premier à montrer les effets toxiques d'une nourriture à base d'un aliment génétiquement modifié, en l'occurrence le maïs NK603 de la firme américaine Monsanto, en association ou non avec du Roundup, l'herbicide auquel il est tolérant. La conclusion des auteurs de l'étude est nette et effrayante : oui, les OGM sont dangereux, et leur consommation peut avoir des conséquences graves pour la santé. Mais est-ce aussi simple et limpide ? Très vite, des critiques ont surgi dans les rangs des scientifiques pour mettre en doute l'étude et ses conclusions.
L'autorité européenne de sécurité des aliments (Efsa) a conclu le 4 octobre 2012 en première analyse que l'étude « présente des lacunes qui ne permettent pas de considérer les conclusions des auteurs comme étant scientifiquement valables ». À l'heure où nous écrivons ces lignes, nous n'avons pas connaissance des conclusions de l'Agence nationale de sécurité sanitaire de l'alimentation, de l'environnement et du travail (Anses), attendues le 20 octobre, mais il est peu probable qu'elles soient clémentes envers les auteurs. Plusieurs points litigieux sont à avoir en tête pour comprendre les débats à venir.
La souche de rats utilisée pour l'expérience, dite Sprague-Dawley, est fragile. Ces rongeurs ont une propension à développer spontanément des tumeurs au cours de leur vie, estimée à deux ans. Une étude de 1973 évalue ainsi l'incidence des tumeurs spontanées à 45 %. De plus, les mâles sont facilement sujets à des atteintes rénales dans des conditions normales.
La composition de l'alimentation des animaux n'est pas détaillée dans l'étude. Or, la nourriture a pu être contaminée par des mycotoxines à l'effet cancérogène et hépatotoxique. De plus les rats Sprague-Dawley développent d'autant plus de tumeurs spontanées que leur nourriture est énergétique et riche en protéines, ce qui est le cas du maïs.
Le faible nombre de rats, dix par groupe, témoins compris, affaiblit la portée statistique de ce travail étant donné qu'il y a très peu de différence, au niveau du nombre et de l'ampleur des pathologies observées entre les rats témoins et ceux nourris aux OGM. Et une étude menée en 2007 au Japon sur du soja tolérant au Roundup avec des effectifs plus importants (50 rats par groupe) n'a rien mis en évidence.
LA RÉPONSE DE GILLES-ÉRIC SÉRALINI
Le chercheur critique les processus d'évaluation des OGM, confiés aux seuls industriels, avec des études courtes (trois mois) et confidentielles. Il dit avoir pris « la même lignée de rats que celle utilisée par Monsanto » et n'empêcher « personne de refaire l'expérience avec 50 rats par groupe ». Il appelle de ses vœux une agence d'évaluation indépendante, financée par l'industrie agroalimentaire.
Vendredi 26 octobre 2012, les Académies nationales d’Agriculture, de Médecine, de Pharmacie, des Sciences, des Technologies, et Vétérinaire ont publié un communiqué au sujet de l'étude de Gilles-Eric Séralini sur l'OGM NK 603 de Monsanto. Leur verdict ? Elle contient "de nombreuses insuffisances de méthodologie et d’interprétation". Paul Deheuvels, lui-même membre de l'Académie des sciences, s'indigne.
Je viens d'apprendre qu'un tout petit nombre de représentants des six Académies (Sciences, Médecine, Technologies,...) se sont réunis pour publier un communiqué commun concernant l'étude du professeur Séralini (éditée récemment dans la revue "Food and Chemical Toxicology").
Les Académies ne sont pas engagées dans leur ensemble
Sans même avoir lu leur déclaration, je me dois d'attirer l'attention du public sur le fait que le dit communiqué ne peut engager l'une ou l'autre de ces académies dans leur ensemble. En effet, un groupe d'experts a été convoqué en urgence, on ne sait par qui, on ne sait comment, dans une absence totale de transparence concernant le choix de ses membres, et sur la base de 2 représentants par académie. Ces personnes ont cru bon de rédiger dans un espace de temps très bref un avis très critique sur cette étude. Elles ne peuvent prétendre à elles seules incarner l'avis de l'ensemble du monde scientifique français, et ce serait une forfaiture que de le laisser croire.
Étant le seul membre de l'Académie des sciences représentant la discipline des statistiques en tant que telle, il aurait été normal que je sois consulté, et tel n'a pas été véritablement le cas.
Il ressort des conversations que j'ai eues a postériori sur ce communiqué que les représentants des cinq académies mentionnés plus haut y aient critiqué la partie descriptive de l'étude du professeur Séralini, concernant les tumeurs, en lui reprochant de ne pas être significative sur le plan statistique. Ils auraient, par contre, ignoré la partie toxicologique de l'article, traitée avec sophistication par des méthodes modernes (dites de PLS). Notons que l'article de Gilles-Eric Séralini est, justement, publié dans une revue de toxicologie.
Un mauvais procès fait à l'étude de Séralini
Dans tout texte du genre, la partie descriptive se contente de décrire, sans pour autant en tirer de conclusion démontrée. C'est bien ce qui se passe ici, et je ne trouve rien à redire sur le plan professionnel à cette composante, quoi que puisse en dire le petit groupe de signataires de la motion.
A l'inverse, la composante de toxicologie de l'article a l'immense mérite d'étudier la cohorte de données complète des analyses réalisée sur une durée d'environ 16 mois. Cette partie de l'article aboutit, quant à elle, à la mise en évidence de différences significatives sur le plan statistique, sous réserve, bien entendu, que leur traitement ait été correctement réalisé, et je ne vois aucune raison de penser que ce n'ait pas été le cas.
On pourra m'objecter que ce sont précisément ces parties descriptives de l'article de Séralini qui ont attiré l'attention des médias, puisqu'elles parlent des tumeurs dont les animaux d'expérience ont été victimes. Ce n'est pas la question. Je pense qu'on fait à cette étude un mauvais procès, par de mauvais arguments, et avec un acharnement parfaitement suspect compte tenu des immenses intérêts financiers qui sont en jeu. Il ne s'agit pas de savoir ce qu'en pensent les médias, mais plutôt de juger la qualité technique de ce travail.
Je tiens à dire, avec force, que l'article du professeur Séralini se situe à un niveau élevé de qualité parmi les articles de même catégorie. On ne peut lui reprocher sa valeur scientifique qui est indéniable. On ne peut l'attaquer sur sa partie descriptive qui ne cherche pas à établir des preuves, au sens statistique du terme, mais à donner une présentation lisible, purement factuelle, des résultats obtenus. Il est d'autant plus anormal que le communiqué des cinq académies ne s'intéresse guère (si mes renseignements sont corrects) à la composante toxicologique de l'analyse, qui, elle, fait usage de niveaux de confiance établis selon les règles habituelles de la statistique.
Pour une saine confrontation des idées
Je récuse donc par avance tout texte qui serait présenté au nom de cinq académies sur ce sujet, partant du fait évident que le comité qui l'a signé ne représente que lui-même, indépendamment de l'éminence de ses membres.
C'est d'ailleurs un procédé à la limite du scandale de vouloir parler au nom de tous lorsqu'on est peu nombreux. Quelle que soit la qualité des signataires, ils expriment un avis qui ne peut prétendre constituer une vérité universelle, tant que le problème n'aura pas été véritablement discuté ouvertement, et sur le fond.
L'article du professeur Séralini a l'immense mérite de mettre en évidence qu'il n'y a pas suffisamment d'études portant sur les effets à long terme des alimentations à base d'OGM. Au lieu de critiquer dans cette étude ce qui ne peut pas l'être, tout en omettant de l'apprécier pour sa composante authentiquement innovatrice, les organismes établis comme l'INRA ou l'ANSES, devraient entreprendre des études approfondies sur des échantillons plus nombreux. Celles-ci permettraient de sortir par le haut d'une querelle aux paramètres évidemment biaisés. Il faut que le débat d'idées ait lieu, dans le calme, et sans avoir à subir les pressions des lobbys qui s'expriment pour ou contre des opinions, sans même prendre le temps de les discuter.
J'exprime un avis personnel, basée sur mon expérience professionnelle, et je n'insulte pas ceux qui ont une opinion contraire. L'intérêt de la science se situe, avant tout, dans une saine confrontation des idées et des arguments, qui soit, si possible, sans parti pris. On est, semble-t-il, très loin d'une telle situation, je le crains.
OGM : Il a montré leur toxicité, il doit être exécuté
Une réaction indignée de Christian Vélot, 19 octobre 2012
L’étude de Gilles-Eric Séralini et de son équipe de l’Université de Caen sur le maïs génétiquement modifié NK603 et l’herbicide Roundup, publiée le 19 septembre 2012 dans l’excellente revue internationale Food & Chemical Toxicology[1], a provoqué un tollé mondial. Il s’agit de l’étude toxicologique la plus longue et la plus complète jamais réalisée au monde sur un organisme génétiquement modifié (OGM) et un herbicide sous sa formulation commerciale : elle a été effectuée sur 200 rats (100 mâles et 100 femelles) pendant deux ans (c’est-à-dire tout au long de leur vie) avec plus de 100 paramètres analysés sur chacun d’eux.
Pour bien comprendre ce qui a conduit G.E. Séralini (GES) et son équipe, soutenus par le CRIIGEN[2] (Comité de recherche et d’information indépendante sur le génie génétique) — dont GES est l’un des fondateurs et le président de son conseil scientifique — à entreprendre une telle étude, et l’acharnement qu’elle suscite, il est nécessaire de faire un bref historique et quelques rappels afin de replacer un certain nombre d’éléments dans leur contexte.
La quasi totalité (99,99%) des OGM agricoles cultivés à la surface de la planète sont des plantes-pesticides. Je rappelle au passage que “pesticide” est le terme générique qui englobe les insecticides (qui tuent les insectes), les fongicides (qui tuent les champignons), les herbicides (qui tuent les herbes, les plantes), etc. Et une plantepesticide est une plante qui accumule dans ses cellules un ou plusieurs pesticides, indépendamment de leur nature. Comment ?
Il s’agit de plantes qui ont été génétiquement modifiées soit pour produire elles-mêmes un insecticide afin de lutter contre un insecte dont elles sont la cible, soit pour ne plus mourir quand on les arrose avec un herbicide qui aurait dû les tuer. Les premières accumulent donc un insecticide parce qu’elles le produisent en permanence, les secondes accumulent un herbicide car elles peuvent l’absorber sans mourir. Alors que la première catégorie correspond donc plus précisément à des plantes-insecticides, la seconde concerne essentiellement des plantes (maïs, soja, colza) qui tolèrent l’herbicide Roundup commercialisé par la société philanthrope Monsanto : c’est le cas du maïs NK603 qui fait donc l’objet de l’étude de GES et de son équipe. Si un agriculteur pulvérise du Roundup sur ses cultures conventionnelles, il va tuer non seulement lesdites “mauvaises herbes”, mais également les cultures elles-mêmes, étant donné que cet herbicide est un herbicide non sélectif qui tuent toutes les plantes (et peut-être pas que les plantes d’ailleurs…). Avec les plantes tolérantes au Roundup, l’agriculteur peut pulvériser l’herbicide en question sans prendre la moindre précaution puisque sa culture tolère l’herbicide : il tuera lesdites “mauvaises herbes” sans détruire la plante cultivée.
Les plantes-insecticides d’une part, et les plantes tolérant le Roundup d’autre part, correspondent aux OGM agricoles dits “de première génération”. Elles représentent, respectivement, environ 20% et 60% de ces OGM, soit 80% au total des plantespesticides. Les 20% manquants (pour atteindre 99,99%) sont les OGM dits de “seconde, troisième et quatrième générations“, c’est-à-dire des plantes génétiquement modifiées qui cumulent plusieurs “caractères pesticides” : la production d’un ou plusieurs insecticide(s) et la tolérance d’un ou plusieurs herbicide(s). On n’arrête pas le progrès…
Les animaux qui vont manger ces plantes vont donc être exposés aux pesticides en question. Les humains qui vont manger les produits animaliers issus des animaux ayant consommé ces plantes vont — éventuellement — êtres confrontés à ces pesticides selon le devenir de ces molécules dans la chaîne alimentaire. La moindre des choses serait donc que ces plantes soient évaluées non seulement comme des plantes, mais également comme des pesticides, ce qui n’est malheureusement pas le cas.
L’évaluation des pesticides utilisés dans l’agroalimentaire est encadrée par la directive européenne 91/414 qui précise qu’ils doivent être soumis à des tests toxicologiques pendant trois mois sur trois espèces animales différentes (dont le rat et la souris, le troisième n’étant pas un rongeur), auxquels s’ajoutent des études à deux ans sur le rat. Pourquoi deux ans? Parce qu’il s’agit justement de la durée de vie d’un rat. Ces études permettent donc d’observer les effets tout au long de la vie, c’est-à-dire les effets chroniques. Alors qu’à trois mois, on ne voit essentiellement que les effets aigus. Cependant, que l’on soit bien clair : loin de moi l’idée selon laquelle ces tests seraient suffisants alors que les pesticides sont connus, dans leur grande majorité, pour se comporter comme des perturbateurs endocriniens, c’est-à-dire pour avoir des effets sur les voies hormonales sexuelles. Il faudrait donc — si l’on s’inscrit dans la démarche de tests sur des animaux — qu’ils fassent également l’objet d’études sur des femelles en gestation, que les descendants soient étudiés sur plusieurs générations, etc. Par ailleurs, les pesticides sont très souvent évalués uniquement sur le principe actif et non sur les formules commerciales qui contiennent un certain nombre de molécules supplémentaires (les adjuvants). Ces mélanges sont pourtant, en général, beaucoup plus dangereux, comme cela a pu notamment être montré à maintes reprises avec l’herbicide Roundup dont les effets toxiques sont bien supérieurs à ceux de son principe actif chez les plantes, le glyphosate. Toutefois, en dépit de toutes ces carences, la législation européenne a au moins le mérite, pour les pesticides, de définir précisément un cadre minimum d’évaluation.
Qu’en est-il des OGM agricoles, c’est-à-dire des plantes-pesticides ? Il s’agit là non plus de la directive 91/414 mais de la directive 2001/18 (qui a été transcrite en droit français début 2008). Et que dit cette directive en matière d’évaluation ? Elle ne dit pas que ces OGM ne doivent pas être évalués, mais elle ne dit pas en quoi doit consister cette évaluation. C’est à l’appréciation des comités d’évaluation, tels que l’ANSES (l’Agence nationale de sécurité sanitaire, de l’alimentation, de l’environnement et du travail) ou le HCB (Haut Conseil des biotechnologies) pour la France, et l’AESA[3] (Agence européenne de sécurité des aliments) pour l’Europe. Dans les faits, les OGM agricoles n’ont jamais été évalués sur le plan toxicologique à plus de trois mois sur une seule espèce animale : le rat (ou la souris). On passe donc systématiquement à côté des éventuels effets chroniques.
Cerise sur le gâteau : les tests sont faits par des laboratoires choisis par les firmes semencières, et pour des raisons de protection industrielle, les données brutes des études en question sont maintenues confidentielles (au sein des agences d’évaluation), excluant ainsi la possibilité de toutes contre-expertises et analyses contradictoires.
Le CRIIGEN n’a eu de cesse de condamner cette carence et cette opacité de l’évaluation des OGM en exigeant d’une part la transparence des données brutes des études à trois mois, et d’autre part des études complémentaires sur deux ans réalisées par des laboratoires indépendants des firmes semencières. G.-E. Séralini a notamment réitéré ces demandes à maintes reprises et en vain au sein de la CGB (Commission du génie biomoléculaire, ancêtre du HCB avant le Grenelle de l’environnement) dans laquelle il a siégé pendant huit années sous la présidence de Marc Fellous, l’un de ses plus fervents détracteurs (qui a d’ailleurs été condamné pour avoir diffamé GES, et qui fait aujourd’hui l’objet d’un second procès de la part de GES et du CRIIGEN pour faux et usage de faux).
Ce n’est qu’au prix de procédures administratives ou judiciaires lourdes que le CRIIGEN a pu – directement ou par l’intermédiaire d’autres organisations — accéder aux données brutes des études à trois mois ayant conduit à l’autorisation de certains OGM de la firme Monsanto : le “fameux” maïs NK 603 et les maïs-insecticide MON 810 (autorisé à la culture commerciale en Europe) et MON 863. GES et le CRIIGEN ont alors réalisé, sur la base de ces données, des contre-expertises publiées en 2007[4] et 2009[5] dans des journaux scientifiques internationaux à comité de lecture[6]. Ces contre-études faisaient apparaître notamment des perturbations du foie et des reins différentes selon les sexes laissant craindre des toxicités hépatorénales chroniques.
Les laboratoires de Monsanto avaient bien observé les perturbations des paramètres hépatiques et rénaux en question, mais avaient conclu qu’elles n’avaient pas de signification biologique, au prétexte qu’elles n’étaient pas identiques dans les deux sexes et qu’elles n’étaient pas non plus proportionnelles à la dose d’OGM contenue dans l’alimentation des différents lots de rats.
On croit rêver ! Les différences selon les sexes laissent évidemment supposer un effet sur les voies hormonales sexuelles. Or ces plantes sont justement des éponges à pesticides qui se comportent très souvent comme des perturbateurs endocriniens. Malgré cela, aucun dosage hormonal n’a été exigé par les agences d’évaluation. Quant à la nonproportionnalité entre les effets et la dose, c’est un phénomène extrêmement courant lorsque les perturbations sont justement d’ordre hormonal. Par exemple, une petite dose de polluants aux propriétés féminisantes peut provoquer l’ovulation, alors qu’une concentration plus importante de ce même polluant peut agir comme une pilule contraceptive, c’est-à-dire de façon totalement opposée.
Peu importe : l’Agence européenne de sécurité des aliments (AESA) a repris à son propre compte les conclusions des laboratoires de Monsanto ("effets biologiquement non significatifs"), et a donné — comme toujours — un avis favorable à la commercialisation et/ou à l’importation pour l’alimentation de ces OGM.
Et les contre-expertises de GES et du CRIIGEN n’y ont rien changé : elles ont été balayées d’un revers de main par l’AESA et les agences nationales (CGB pour l’étude de 2007 ou HCB pour l’étude de 2009) qui ont conclu, en utilisant les mêmes arguments, que ces études n’étaient pas de nature à remettre en question l’innocuité des OGM en question. Circulez, il n’y a rien à voir !
Mais c’était sans compter sur la ténacité de GES. Soutenu par le CRIIGEN, il a alors entrepris de collecter les fonds (3 millions d’euros) dans le but de réaliser lui-même avec son équipe, et en s’entourant des collaborateurs apportant les différentes compétences nécessaires, l’étude à deux ans sur l’un de ces maïs : le NK 603.
Dans cette étude, GES ne s’est pas contenté d’étudier les effets de l’OGM (à trois doses au lieu de 2 dans toutes les autres études), mais également du Roundup (à trois doses également) auquel ce maïs a été rendu tolérant, ainsi que de la combinaison des deux (OGM + Roundup). Le but étant de savoir si les éventuels effets de cet OGM étaient dus à l’OGM lui-même ou aux résidus de Roundup qu’il accumule (ou aux deux). Dans chaque sexe, les 100 rats ont donc été divisés en 10 lots de 10 : un lot contrôle (qui ne mange pas d’OGM et ne boit pas de Roundup), 3 lots nourris avec OGM (à trois doses différentes), trois lots traités au Roundup (à trois doses différentes) et trois lots traités à la fois à l’OGM et au Roundup.
Les résultats révèlent non seulement, comme on pouvait s’y attendre au regard des contre-expertises précédentes, des effets hormonaux non linéaires et reliés au sexe, mais également des mortalités plus rapides et plus fortes au cours de la consommation de chacun des deux produits. Les femelles développent des tumeurs mammaires importantes et en grand nombre, ainsi que des perturbations hypophysaires et rénales, et les mâles présentent des atteintes hépatorénales chroniques sévères.
Évidemment, les réactions n’ont pas tardé. Dès le premier jour après sa publication, l’étude a fait l’objet d’attaques violentes, émanant essentiellement, comme par hasard, de ceux qui sont à l’origine de l’autorisation de ce maïs, tels Marc Fellous et Gérard Pascal, et ce avant même qu’ils aient pu la lire en détail et s’en imprégner. On comprend qu’ils soient piqués au vif, cette nouvelle étude remettant directement en cause la validité de leur propre évaluation. Et ceux là-mêmes qui se sont toujours opposés aux contre-expertises, demandées entre autres par GES et le CRIIGEN, deviennent tout d’un coup les fervents défenseur des contre-analyses et de l’expertise contradictoire : "cette étude doit être validée par des experts", des vrais, qui représentent LA communauté scientifique. Ben voyons. Je rappelle quand même que l’étude de GES n’a pas été publiée dans Pif le chien mais dans un journal à comité de lecture (qui plus est, très coté dans le monde de la toxicologie), ce qui signifie qu’elle a déjà été expertisée par les pairs (par des experts internationaux choisis anonymement par le bureau éditorial du journal).
Bien sûr, ceci n’exclut pas la critique. Il n’existe pas d’étude scientifique, et quelle que soit la notoriété du journal dans lequel elle est publiée, qui soit irréprochable et qui ne puisse faire l’objet de critiques. La contradiction, c’est ce qui fait respirer la démocratie. On en a besoin en sciences comme ailleurs. C’est sain. Mais ces critiques doivent être faites de façon constructive, selon certaines règles de déontologie, dans le but d’identifier les éventuels points faibles et proposer en fonction les modifications de protocole nécessaires à la répétition de l’étude tout en s’affranchissant d’éventuels obstacles ou certaines difficultés. Le mot clé en sciences est “reproductibilité”. Cette étude étant une première, elle doit de toute façon être refaite. Avec comme seule préoccupation la sécurité sanitaire et non pas le flinguage d’un collègue trop gênant. Voilà ce qu’on est en droit d’attendre de scientifiques dignes de ce nom.
Au lieu de cela, on assiste à un lynchage en règle de Séralini et ses co-auteurs. Leur étude ne vaut pas un pet de lapin. Tout est à jeter. Certes, ces attaques viennent essentiellement d’un petit groupe de scientifiques constitué des militants pro-OGM habituels — les Felllous, Pascal, Dattée et autres Joudrier de l’AFBV[7] (Association française des biotechnologies végétales) —, mais qui a le culot de prétendre représenter LA communauté scientifique dans une tribune largement relayée par le journal Marianne et son journaliste expert en OGM, Jean-Claude Jaillette. Et toujours avec les mêmes arguments stupides utilisés qui tournent en boucle depuis le premier jour.
1) Les rats utilisés ne sont pas les bons car ils sont réputés très sensibles aux cancers. Ah bon ? Mais ce sont pourtant ceux utilisés dans toutes les études de Monsanto qui ont conduit, entre autres, à l’autorisation de ce maïs. Et ce sont aussi ceux qui sont utilisés dans toutes les études de toxicologie, qu’il s’agisse de médicaments ou de pesticides.
En fait, quand on ne voit rien, ces rats sont parfaits, quand on voit des effets, c’est que les rats choisis ne sont pas les bons. Chat perché ! Sans doute Séralini et ses collaborateurs auraient-ils dû choisir des rats résistants aux cancers pour ne pas offusquer tous ces scientifiques bien pensants et, pour certains, contredire leur propre expertise. Vous voulez étudier le cancer de la prostate ? Faites-le sur des femmes, c’est plus sûr.
2) Les effectifs de rats traités et contrôles (lots de 10 rats) sont insuffisants. Ah bon ? Alors, pourquoi n’a t-on pas entendu ces scientifiques exemplaires protester contre toutes les études ayant conduit aux autorisations des OGM et pour lesquelles les groupes de rats n’ont jamais excédé 10 ? Pour la pomme de terre transgénique Amflora, il s’agissait même de groupes de 5 rats, et pour le maïs MIR 162 qui est sur le point d’être autorisé, des groupes de 6 rats.
En fait, quand on ne voit rien, 5 rats ça suffit, quand on voit des effets, 10 rats, c’est insuffisant et donc l’étude ne vaut rien. Chat perché ! Vous me suivez ? Oui, je sais, c’est compliqué la science !
3) Cette étude n’est pas la première étude de long terme effectuée pour des OGM. Il y en a eu beaucoup d’autres faites sur d’autres animaux, notamment sur des porcs pendant trois ans. Et aucun effet n’a jamais été observé ? Alors là, nos scientifiques de référence deviendraient-ils de mauvaise foi ou feraient-ils preuve d’une certaine incompétence ? Rappelons d’abord que deux ans pour des rats, c’est la vie entière. Ce qui n’est pas le cas de trois années pour le porc qui vit au moins quinze ans. Par ailleurs, les études auxquelles il est fait référence ne sont en rien des études toxicologiques, mais des études nutritionnelles qui consistent simplement à nourrir l’animal et à observer quelques paramètres macroscopiques (poids, volume de lait produit,…) sans faire la moindre analyse de sang, la moindre analyse d’urine, la moindre biopsie ou prélèvement d’organe, le moindre dosage hormonal, la moindre palpation pour détecter d’éventuelles grosseurs ou tumeurs, etc.
4) Des millions d’animaux et d’américains mangent des OGM tous les jours depuis des années et on n’a jamais rien observé de la sorte. Cet “argument” n’est pas nouveau. C’est notamment celui qu’avait utilisé Axel Kahn lors d’une émission « La tête au carré » sur France Inter. A la question d’un auditeur relayée par l’animateur Mathieu Vidard, demandant à l’invité s’il trouve normal que les OGM agricoles ne soient pas testés plus de 90 jours sur quelques dizaines de rats, et s’il pense vraiment que des tests aussi réduits peuvent assurer une innocuité raisonnable de ces produits, A. Kahn répond que "nous disposons d’un test en grandeur nature, à savoir qu’il y a 200 millions d’Américains qui mangent des OGMs depuis 10 à 20 ans et qu’aucun problème de santé n’a été recensé".
Si de tels propos avaient été tenus par Claude Allègre, nous aurions compris. Mais enfin, Axel Kahn, le généticien référent du journal de 20H ! Comment est-ce possible ? Sur quelles études sanitaires à grande échelle se base-t-il pour tenir de telles affirmations ? Sachant que depuis que les OGM existent sur le continent américain, il n’y a jamais eu aucun suivi systématique des animaux et des humains qui en ont mangé. Et comment de telles études pourraient-elles d’ailleurs exister dans la mesure où ce continent ne sépare pas les filières agricoles? Il est donc impossible de savoir qui consomme des OGM, à quelle dose et à quelle fréquence, et qui n’en consomme pas. Comment pourrait-on alors établir une corrélation de cause à effet entre l’apparition d’un quelconque problème sanitaire et la consommation d’OGM ? D’autant plus que les effets montrés par GES et son équipe sont justement des effets chroniques, qui peuvent donc, au sein d’une population, être masqués par d’autres effets chroniques aux origines multiples M. Kahn ignore t-il lui aussi ce qu’est une étude toxicologique ? Décidément, la science n’est plus ce qu’elle fut.
Cet argumentaire de haute volée a été repris dans un article paru sur Médiapart le 22 septembre dernier. Son auteur, Michel de Pracontal, semble s’y connaître autant en OGM que Nadine Morano en communication, au point qu’il n’a toujours pas compris le lien entre le maïs NK603 et le Roundup., et que son texte est cousu d’inepties à faire rougir Claude Allègre.
Nos scientifiques exemplaires s’offusquent également du "coup médiatique" qui a accompagné la sortie de cette étude: "Une telle démarche n’est pas une démarche scientifique éthiquement correcte". Bien sûr, ils auraient tellement préféré que cette étude reste dans le cercle fermé des scientifiques savants qui savent et que surtout les bouseux incultes, les sans grade et les gueux incapables de comprendre n’y aient pas accès. C’est une affaire de scientifiques enfin ! Ne mélangeons pas tout ! Et bien non, désolés de vous décevoir messieurs les référents en matière d’éthique, c’est avant tout une affaire qui concerne l’ensemble de la société. Chacun a le droit de savoir avec quelle transparence et quelle rigueur sont évalués les produits qui se retrouvent dans son assiette. Messieurs les scientifiques éthiquement responsables considèrent-ils que l’embargo sur les données brutes des études ayant conduit aux autorisations des OGM et des pesticides, et le fait qu’elles soient réalisées par leurs promoteurs, soient une démarche éthiquement correcte ? Ils sont soucieux de l’éthique depuis quand, ces messieurs de LA communauté scientifique : depuis le 19 septembre 2012 ? Si les méthodes de communication utilisées par GES et le CRIIGEN ont permis un déclic éthique chez certains de nos collègues, c’est déjà ça de gagné. On avance.
C’est ensuite au tour de l’agence européenne de sécurité alimentaire (AESA) – dont on connaît l’impartialité puisqu’elle a elle-même autorisé les produits évalués par GES et son équipe en émettant, comme toujours, un avis favorable auprès de la Commission européenne — et de l’Agence allemande de sécurité sanitaire BfR de donner leur avis sur l’étude. Et les conclusions des deux agences sont quasiment les mêmes au mot près : étude pas suffisamment robuste pour être validée. Et encore une fois sur la base des mêmes “arguments” foireux que ceux exposés ci-dessus et exploités par nos experts de l’éthique.
Ces deux agences ont réussi l’exploit d’évaluer en moins de deux semaines l’étude qui a nécessité 4 mois d’expertise de la part des experts internationaux de la revue Food & Chemical Toxicology. Et pour justifier un tel exploit, M. Jaillette, qui s’empresse de rapporter dans Marianne les conclusions de l’Agence BfR, précise qu’[elle] a mis les bouchées doubles (sic). Elle a tellement mis les bouchées doubles qu’elle n’a visiblement pas bien digéré l’étude et qu’elle n’y a, en tout cas, pas compris grand chose. L’agence BfR précise notamment que [cette étude] n’apporte aucune raison valable pour rééxaminer le glyphosate. Alors qu’elle concerne non pas le glyphosate mais le Roundup dans sa formule commerciale qui, lui, n’a jamais été évalué en tant que tel. GES n’a jamais demandé une réévaluation du glyphosate mais une évaluation du Roundup. Et Jaillette, en plein orgasme, qui s’empresse de conclure sur la base de ces analyses oh combien pertinentes : "Voici donc l’étude Séralini taillée en pièce…". Jaillette en a rêvé, l’Agence BfR l’a fait.
Et la furie continue : c’est tout récemment les académies scientifiques qui s’en mêlent en rendant publique une analyse très sévère de l’étude de GES. Alors là, évidemment, même si les académies le disent… Doit-on rappeler l’avis de l’académie de médecine sur l’amiante qui en minimisait les risques et confortait les thèses du lobby de ce poison[8] ? Doit-on rappeler l’avis de l’académie des sciences sur les dioxines qui en minimisait la toxicité et concluait que ces dernières ne constituaient pas un risque majeur pour la santé publique[9] ? On peut donc leur faire confiance…
Et non contentes de vilipender l’étude de GES, les académies, dans leur avis, vont jusqu’à recommander la création d’un "Haut comité de la science et de la technologie" dont la mission serait d’attirer l’attention du CSA (Conseil supérieur de l’audiovisuel) sur la médiatisation de travaux scientifiques remettant en cause des savoirs partagés par la très grande majorité de la communauté scientifique internationale. Magnifique ! Vive le retour au régime de Vichy ! Un comité de censure pour étouffer toutes les études scientifiques qui remettent en cause les dogmes et la pensée dominante de LA communauté scientifique ! On n’y avait pas pensé ! Mais qui a dit que les académies scientifiques ne servaient à rien ?
Face à un tel acharnement, et malgré le fait qu’il soit soutenu par de nombreux scientifiques en France et à travers le Monde, Gilles-Eric Séralini se retrouve dans la très inconfortable position du lanceur d’alerte, en proie aux représailles d’industriels menacés ou d’experts autoproclamés.
L’absence actuelle, en France notamment, de dispositif juridique permettant une protection du lanceur d’alerte et un encadrement du traitement de l’alerte est d’autant plus tragique que la recherche et son développement sont pilotés de plus en plus largement par des intérêts industriels à court terme nécessitant des retours sur investissements rapides. Cette politique — qui est le résultat de la déresponsabilisation progressive, depuis plusieurs décennies, des gouvernements successifs en matière de financement de la recherche publique — conduit inévitablement à de graves dérives, et notamment, comme c’est typiquement le cas avec les OGM, à une carence et une opacité des évaluations sanitaire et environnementale, le temps que demandent ces évaluations n’étant pas compatible avec l’urgence des brevets et des profits d’une part, et la protection industrielle justifiant l’absence de communication des données brutes issues des analyses des risques d’autre part. Dans un tel contexte, l’alerte et l’expertise prennent donc une place tout à fait primordiale. Il est donc urgent que la France se dote, dans son arsenal juridique, d’un dispositif de protection du lanceur d’alerte environnementale et sanitaire, à l’instar de pays comme les États-Unis et leur "Whistleblower Protection Act", ou comme la Grande-Bretagne ("Public Interest Disclosure Act"). Mais pour être réellement efficace, un tel dispositif juridique doit se replacer dans un contexte de loi réformant le système de l’expertise et reposant sur le principe de précaution.
Le 15 octobre 2012, le groupe écologiste du Sénat a présenté en première lecture une proposition de loi sur la protection des lanceurs d’alerte et la déontologie de l’expertise scientifique dans le domaine de la santé et de l’environnement, prévoyant notamment la création d’une Haute Autorité de l’expertise scientifique et de l’alerte qui serait chargée d’énoncer les principes directeurs de l’expertise scientifique et technique en matière de santé publique et d’environnement, d’en vérifier l’application et de garantir la mise en oeuvre des procédures d’alerte. Cette proposition, portée par la sénatrice du Nord, Marie-Christine Blandin, a été largement inspirée par les travaux de la Fondation Sciences Citoyennes[10] qui réclame depuis plusieurs années une telle avancée législative, ainsi que par le rapport d’étapes des travaux de la Commission Lepage sur la gouvernance suite au Grenelle de l’environnement. Les débats sur la mise en place d’une telle loi devraient reprendre au Sénat le 21 novembre 2012.
Le retentissement médiatique de certaines affaires, comme celles du Médiator avec Irène Frachon ou des OGM avec Gilles-Eric Séralini nous montre à quel point il est urgent de réformer totalement les systèmes d’expertise actuels, trop souvent gangrenés par l’opacité et les conflits d’intérêt, afin que les problématiques de santé et d’environnement ne soient plus à la merci des intérêts mercantiles, et que les quelques scientifiques qui ont le courage d’alerter en dénonçant et démontrant les carences d’une évaluation ne soient pas envoyés à l’échafaud.
Christian Vélot, Généticien moléculaire à l’Université Paris-Sud 11
Courriel : christian.velot@u-psud.fr
Membre du Conseil scientifique du CRIIGEN
Administrateur de la Fondation Sciences Citoyennes
Auteur de « OGM, tout s’explique » (Eds Goutte de Sable, 2009) et « OGM, un choix de société » (Eds de l’Aube, 2011)
[1] G.E. Séralini, E. Clair, R. Mesnage, S. Gress, N. Defarge, M. Malatesta, D. Hennequin, J. Spiroux de Venômois. Long term toxicity of a Roundup herbicide and a Roundup-tolerant genetically modified maize. 2012. Food and Chemical Toxicology 50 : 4221-4231
[2] Cofondé par Corinne Lepage, Gilles-Eric Séralini et Jean-Marie Pelt. http://www.criigen.org/
[3] Elle est parfois plus connue sous l’acronyme anglais : EFSA (European Food Safety Authorithy)
[4] G.E. Séralini, D. Cellier, J. Spiroux de Vendômois. New Analysis of a Rat Feeding Study with a Genetically Modified Maize Reveals Signs of Hepatorenal Toxicity. 2007. Archives of Environmental Contamination and Toxicology 52 : 596-602
[5] J. Spiroux de Vendômois, F. Roullier, D. Cellier, G.E. Séralini. A comparison of the effects of three GM corn varieties on mammalian health. 2009. International Journal of Biological Sciences 10 : 706-726
[6] Un journal à comité de lecture est un journal où toute publication scientifique nécessite une expertise préalable du contenu de l’étude par des experts internationaux choisis anonymement par le bureau éditorial
[7] Association de scientifiques agissant comme groupe de pression en faveur des OGM agricoles en se masquant derrière une prétendue neutralité scientifique
[8] http://www.liberation.fr/sciences/0101227124-amiante-le-rapport-qui-fache-la-publication-desconclusions-de-l-inserm-s-est-heurtee-a-une-serie-de-blocages
[10] La Fondation Sciences Citoyennes a pour objectif de favoriser et prolonger le mouvement actuel de réappropriation citoyenne et démocratique de la science, afin de la mettre au service du bien commun. http://sciencescitoyennes.org/.
Concernant toujours la publication de G.-E. Séralini, le magazine "Pour la Science" de novembre 2012, page 6-7 propose un article de Loïc Mangin : OGM : une étude qui en appelle d'autres.
Le même magazine "Pour la Science" de janvier 2013 (n° 423), annonce que l'étude Séralini est rejetée par les instances officielles :
"Publiée le 19 septembre 2013, dernier dans une revue internationale, l'étude de Gilles-Éric Séralini, de l'Université de Caen et membre du CRIIGEN (un comité de recherche indépendant), a déchaîné les passions en France. Réalisée sur des rats, elle affirmait démontrer la toxicité du maïs transgénique NK603 vendu par Monsanto et de l'herbicide Roundup auquel cet OGM est résistant. Les passions et l'enjeu étaient tels, que plusieurs instances officielles ont dû se pencher sur ces travaux et se prononcer sur leur validité. Avec un verdict négatif : l'ANSES (Agence nationale de sécurité sanitaire de l'alimentation, de l'environnement et du travail), le HCB (Haut Conseil des biotechnologies), l'EFSA (Autorité européenne de sécurité des aliments) ont tous trois disqualifié l'étude Séralini (échantillon statistiquement trop petit, souche de rats inadaptée à des études sur deux ans...). Mais l'affaire aura au moins permis de souligner la nécessité, dans ce domaine sensible, de travaux indépendants, transparents et rigoureux - y compris du côté des industriels!"
http://www.criigen.org, http://www.anses.fr, http://www.hautconseildesbiotechnologies.fr, http://www.efsa.europa.eu/fr/
Quoiqu'on en pense, les travaux de G.-E. Séralini auront réveillé les consciences et la Commission européenne. Voir l'article du magazine "Sciences et Avenir".
OGM : les institutions contre l'étude Séralini
Rachel Mulot (Science et Avenir n° 790, décembre 2012. p. 40)
Bien qu'"ambitieuse et originale", l'étude sur la toxicité des OGM par le professeur Gilles-Eric Séralini[1] échoue à apporter la preuve d'un lien entre la consommation de maïs NK603 et l'apparition de pathologies chez les rats nourris pendant deux ans avec cet OGM, traité ou non avec l'herbicide Roundup (voir S. et A. n° 789, novembre 2012, et le site Internet de Sciences et Avenir).

Tel est l'avis de l'Anses (Agence nationale de sécurité sanitaire), qui, saisie par le gouvernement, a détaillé les faiblesses méthodologiques et statistiques de l'étude[2]. Le Haut Conseil des biotechnologies (HCB) a repoussé les conclusions de l'article de la même façon[3]. Toutefois, les deux organismes publics recommandent que soient désormais menées, de façon indépendante, des recherches sur les effets à long terme des OGM associés aux pesticides. Seules trois études portant sur le rat, dont celle du Pr Séralini, ont été a ce jour été consacrées à ces effets à long terme. Et elles ne permettent pas de conclure sur le sujet.
Dans la foulée de ces avis, le gouvernement a plaidé pour une " remise à plat du dispositif européen d'évaluation, d'autorisation et de contrôle des OGM et des pesticides ». En outre, quatre ex-ministres de l'Environnement ont demandé que soient revues, de façon indépendante, et avec la même sévérité critique, les études qui ont permis l'autorisation du maïs NK603 et du Roundup[4].
Autant de révisions que l'Efsa, l'organisme européen en charge des OGM, avait estimé inutiles...
En attendant, Gilles-Éric Séralini s'apprêterait à publier, toujours dans la revue Food and Chemical Toxicology des articles complémentaires afin de répondre aux questions et aux critiques. R. M.
"Ce qu'on sait vraiment des OGM"
C'est le titre du dossier ouvert par Rachel Mulot et Hervé Ratel dans le magazine Sciences et Avenir n° 791 de janvier 2013 qui fait le bilan pages 8 à 13 des études effectuées sur l'innocuité des OGM en exposant les arguments des pro-OGM, des anti-OGM et des agences comme l'ANses ou le HCB qui se situent entre deux-feux. Ce dossier conclut sur la nécessité de financer des études à long terme menées par des organismes réellement indépendants. Voir également l'article publié sur le site de Sciences et Avenir.
22:43 Publié dans Biologie, Botanique, Environnement-Écologie, Pollution | Tags : ogm, séralini, cancer | Lien permanent | Commentaires (2) | Facebook | | |
28/07/2012
Des cellules souches dans la rétine
 Des cellules souches dans la rétine
Des cellules souches dans la rétine
(Dernière mise à jour : 2/11/2014)
Dans ce même blog, nous avons évoqué l'emploi de cellules souches embryonnaires pour régénérer la rétine. Ces cellules embryonnaires sont pluripotentes car elles sont à l'origine de toutes les cellules de l'organisme (voir dans ce même blog les articles : Cellules souches et rajeunissement et Élaboration d'une rétine à partir de cellules souches embryonnaires).
Une autre technique est utilisée dans le même but de régénération de la rétine par une autre équipe américaine : des cellules souches découvertes dans la rétine d'individus adultes décédés ouvrent une nouvelle piste de traitement de l’une des premières causes de cécité, la dégénérescence maculaire liée à l’âge, ou DMLA.
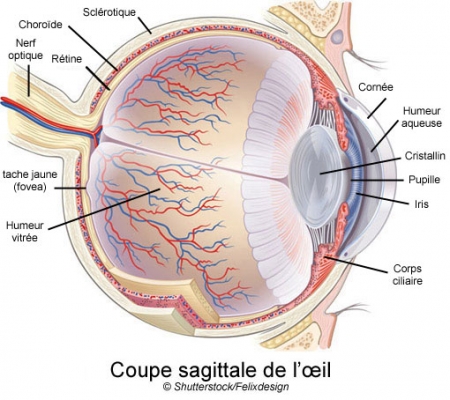
La dégénérescence maculaire liée à l'âge (DMLA) détruit les cellules de l'épithélium pigmentaire de la rétine. Cette couche de cellules qui tapisse le fond de la rétine assure le bon fonctionnement des cellules photoréceptrices de l'œil. Une maladie qui n'est pas négligeable et qui s'aggrave avec l'âge des patients. D'après le journal d'ophtalmologie, 2007, le taux de DMLA dans le monde, s'établit selon l'âge : de 55 à 64 ans à 12,2%, 65 à 74 ans à 18,3%, plus de 74 ans à 29,7% (600 000 patients en France).
Les travaux de l’équipe de Sally Temple, de l’Institut des cellules souches neuronales de Rensselaer, aux États-Unis ouvrent un espoir de remédier à la DMLA. En cultivant sous diverses conditions de telles cellules prélevées sur des donneurs décédés, les biologistes ont mis en évidence, parmi elles, l’existence d’une sous-population de cellules souches multipotentes, c’est-à-dire capables de se différencier en plusieurs types cellulaires – dont les cellules de l’épithélium pigmentaire de la rétine.

La DMLA (dégénérescence maculaire liée à l'âge) entraîne l'apparition d'une ou plusieurs taches noires qui occultent la vision centrale
Cet épithélium est une couche de cellules qui tapisse le fond de la rétine et assure le bon fonctionnement des cellules photoréceptrices de l’œil, les bâtonnets et les cônes. La DMLA, principale cause de malvoyance et de cécité dans les pays industrialisés – elle y touche sept pour cent des personnes de plus de 75 ans –, est due à la disparition progressive de ces cellules. Il n’existe encore aucun traitement pour la forme la plus fréquente, dite DMLA « sèche » ou atrophique.
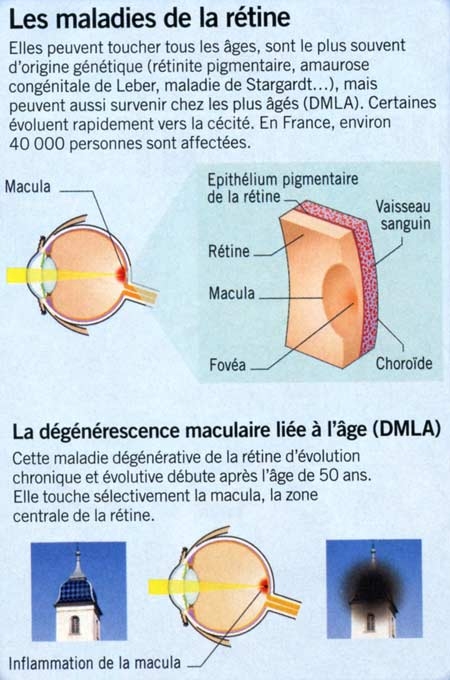
Infographie : Betty Lafon pour Sciences et Avenir
Une piste explorée par plusieurs équipes consiste à greffer dans la rétine des cellules de l’épithélium pigmentaire de la rétine produites en laboratoire à partir de cellules souches embryonnaires (cellules ES) ou de cellules induites à la pluripotence (cellules iPS), c’est-à-dire capables de se différencier en cellules de n'importe lequel des trois feuillets embryonnaires (lors de l’embryogenèse, ces feuillets produisent les organes du futur individu).
La DMLA (dégénérescence maculaire liée à l'âge) se caractérise par une atteinte de la zone centrale de la rétine, la macula (le cercle jaune vu en rétino-angiographie)
© KOKEL/BSIP
En découvrant des cellules souches dans l’épithélium pigmentaire de la rétine humaine adulte, les biologistes américains ouvrent la voie à une nouvelle piste : en culture, sous certaines conditions, ces cellules souches multipotentes se différencient en cellules de l’épithélium pigmentaire. Existerait-il des conditions similaires qui activeraient leur différenciation dans la rétine des personnes atteintes de la maladie, et ainsi la régénération in situ de l’épithélium défectueux ? Certains animaux à sang froid, comme les amphibiens, ont déjà cette capacité de régénérer leur épithélium pigmentaire et d’autres tissus oculaires lorsque ceux-ci sont abîmés. Prochaine étape : décortiquer le mécanisme qui active la différenciation de ces nouvelles cellules souches – une donnée indispensable pour mettre au point les conditions favorables à la régénération de l’épithélium in situ.
« C’est la première fois que l’on met en évidence des cellules souches dans ce tissu chez l’adulte, commente Olivier Goureau, de l’Institut de la vision, à Paris. Cette piste de réparation endogène semble prometteuse. » Celle de la greffe de cellules de l’épithélium pigmentaire dérivées de cellules pluripotentes n’est pas abandonnée pour autant. Son équipe, qui travaille sur la production de différentes cellules rétiniennes à partir de cellules souches pluripotentes humaines, s’apprête, en collaboration avec le laboratoire ISTEM du génopole d’Evry, à effectuer chez la souris des tests de greffe d’un tel épithélium.
Pour en savoir plus
E. Salero et al., Adult human RPE can be activated into a multipotent stem cell that produces mesenchymal derivatives, Cell Stem Cell, vol. 10, pp. 88-95, 2012.
C. Delcourt, Les troubles de la vision liés à l'âge, Pour la Science, n° 292, pp. 38-45, 2002.
Les cellules souches embryonnaires humaines tiennent leurs promesses (ajout de janvier 2013)
Des patients ont reçu une greffe de cellules souches embryonnaires humaines. Une première mondiale qui offre un espoir de traitement pour cette maladie incurable.
De gauche à droite, tomographie par cohérence optique de la rétine d'une macula normale, d'une rétine atteinte de DMLA et enfin d'un œdème maculaire, une complication rencontrée souvent chez les diabétiques
(CMCR MAISON ROUGE STRASBOURG)
Pour la première fois, une équipe du prestigieux Jules Stein Eye Institute de Los Angeles (États-Unis) a réalisé avec succès, en mars 2012, une greffe de cellules souches embryonnaires humaines (CSEh) chez deux patientes atteintes de dégénérescence maculaire liée à l'âge (DMLA). L'une des patientes souffrait de la forme dite sèche. L'autre d'une affection proche appelée maladie de Stargardt. Ces CSEh, prélevées sur l'embryon aux premiers stades de développement, ont la particularité de se répliquer indéfiniment avant de se différencier pour former plus de 200 types de tissus humains. 50 000 d'entre elles ont été injectées sous la rétine de patientes très malvoyantes pour reconstituer de la rétine. Six mois après, l'une pouvait à nouveau se déplacer seule, l'autre était capable d'utiliser partiellement un ordinateur. Un succès. Mais ces résultats doivent, bien sûr, être confirmés sur la durée. C'est pourquoi l'étude va se poursuivre pendant encore deux ans et inclure vingt-quatre autres patients.
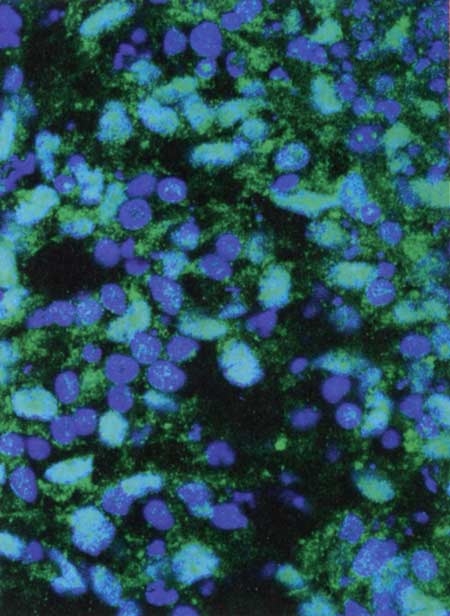
Les greffes de cellules souches embryonnaires humaines (ci-dessus, vues au microscope) sont peut-être la thérapie de l'avenir. Au dernier stade de leur développement, elles donneront un véritable tissu rétinien, aidant à restaurer la vision du patient
© UC IRVINE
S'ils sont avérés, ces résultats constitueront un immense espoir de traitement pour les deux formes de cette maladie dont les symptômes sont une déformation des lignes droites avec l'apparition progressive d'une ou plusieurs taches noires centrales, les scotomes, traduction de la perte de la vision centrale. Moins fréquente (20 % des cas), la forme dite humide est la plus grave. Elle se caractérise par la formation de néovaisseaux sous la rétine, une prolifération due à une production anormale de facteurs de croissance (VEGF : Vascular Endothelial growth factor). En revanche, il n'existe à ce jour aucun traitement pour la forme sèche sans néovascularisation (dite aussi atrophique, plus lente et plus répandue (80 %), les antiVEGF étant inopérants. L'objectif thérapeutique est dans ce cas de stabiliser la destruction progressive des cellules photoréceptrices de la rétine et de bloquer l'inflammation locale. « Plusieurs molécules sont à l'étude et des essais cliniques vont démarrer dans les mois qui viennent », prévoit le Pr Eric Souied, président de l'Association DMLA*.
Face à une affection dont le nombre de cas pourrait doubler d'ici à 2030 en raison du vieillissement de la population, les chercheurs s'intéressent aussi au dépistage. On sait ainsi de longue date que le risque de développer une DMLA est plus important en cas de tabagisme et de surpoids. Plus récemment, des travaux ont été consacrés à la prédisposition génétique et plusieurs marqueurs ont été identifiés (ARMS2, CFH...). Un test de dépistage à partir d'un simple prélèvement salivaire, Macula Risk, est même déjà fabriqué et commercialisé au Canada et sur Internet par la firme Artic DX. Toutefois, son usage reste controversé et surtout prématuré comme l'a précisé un communiqué spécial émis par l'American Academy of Ophtalmology (AAO) en novembre 2012. « À l'avenir, ces tests pourront probablement aider à mieux traiter les patients à haut risque en permettant des traitements adaptés, a ainsi précisé le Dr Edwin Stone, responsable du groupe de travail de l'AAO sur la prédisposition génétique. « Mais les preuves d'un réel bénéfice pour le patient manquent encore. » Pour l'heure, en France, ce test est depuis peu en cours d'évaluation au centre hospitalier intercommunal de Créteil (Val-de-Marne) selon un protocole rigoureux, et uniquement, auprès de patients présentant des antécédents familiaux. L'équipe de Créteil poursuit par ailleurs un projet de génétique appliquée. « Nous avons identifié des marqueurs génétiques prédictifs de sévérité, détaille le Pr Souied. D'ici à cinq ans, nous espérons pouvoir adapter notre prise en charge au profil de chacun. » Un premier pas de soin personnalisé de la DMLA.
*N° vert 0.800.880.66.
Source :
Sylvie Riou-Millot (2013) . - Bien voir - Sciences et Avenir n° 792, février 2013, pp. 46-57.
Les cellules souches induites adultes (iPS) entrent dans la danse (ajout de mars 2013)
Dans le traitement de la DMLA, les premiers essais cliniques impliquant des cellules iPS devraient démarrer au Japon en 2013 au centre hospitalier de Kobe. Cette opération, qui se pratique déjà avec les CSEh (voir plus haut et également le dossier « Vision », Sciences et Avenir n° 792, février 2013) consistera à injecter des iPS différenciées directement dans l'œil du patient afin de reconstituer les cellules abîmées.
Peut-on envisager de déployer cette technique pour des interventions nécessitant de plus importantes réparations dans des tissus nécrosés ? Dans le cas de la rétine, le nombre de cellules iPS à injecter est relativement faible. Mais s'il est possible d'envisager de greffer en surface de l'œil quelques milliers de cellules rétiniennes au millimètre carré, sera-t-il possible de réparer en profondeur un trouble cardiaque comme l'ischémie qui détruit plusieurs dizaines de milliards de cellules ? Sera t-il possible de produire autant de cellules, et à quel coût ? Autant de questions qui restent sans réponse. Pour toutes ces raisons, il semble illusoire - du moins pour l'heure - d'envisager que les iPS puissent ouvrir la voie à une médecine totalement personnalisée où l'on pourra soigner chaque patient à partir de ses propres cellules expurgées de leurs défauts.
Une greffe relance l'espoir dans la DMLA (ajout du 2 novembre 2014)
Une première mondiale : l'équipe menée par Yasuo Kurimoto, du Kobe City Médical Center Général Hospital au Japon a réalisé un implant de tissu rétinien à partir de cellules souches pluripotentes induites (iPS).
Le 12 septembre 2014, une Japonaise de 70 ans atteinte de DMLA a reçu une greffe d'un tissu rétinien conçu à partir de cellules souches pluripotentes induites (IPS). Une première mondiale.
Même s'il est trop tôt pour évaluer le résultat, la patiente a déclaré, le lendemain, que sa vue était plus "brillante". Les iPS en question étaient des cellules souches obtenues à partir de cellules adultes différenciées — ici de peau de la patiente — puis reprogrammées en cellules de rétine. L'équipe japonaise a implanté un feuillet de 1,3 sur 3 mm d'épithélium rétinien dans l'œil de la patiente.
« Cette excellente équipe a réalisé ce que nous appelons "une preuve de concept" », commente Marc Peschanski, directeur scientifique d'I-Stem au génopôle d'Évry (Essonne). Nous attendons les résultats dans les deux ans. » Un éventuel succès pourrait cependant être difficile à étendre au plus grand nombre. « Le procédé n'étant pas du tout industrialisé, nous allons nous retrouver devant un problème éthique : nous aurons le traitement mais pas les moyens de l'appliquer. »
Source : Article de Sciences et Avenir, n° 813, novembre 2014 p. 28.
Une piste autre que les cellules souches : Un collyre anti-cholestérol pour ralentir la DMLA
Une possibilité de ralentir la maladie vient d'être explorée par une équipe de chercheurs américains dirigée par Rajendra Apte, du département d'ophtalmologie de l'École de médecine de l'université Washington à Saint-Louis (Missouri).
Principale cause de malvoyance chez les personnes âgées, la DMLA pourrait être prévenue par des gouttes oculaires évacuant le cholestérol de l'œil. Un essai prometteur chez le rat.
Les injections dans l'œil utilisées pour traiter localement la DMLA pourraient céder la place à un simple collyre. C'est en effet la piste ouverte par des chercheurs américains qui publient une étude dans Cell Metabolism. Selon ce travail, il serait possible de ralentir la prolifération anormale des vaisseaux qui caractérise cette affection en régulant le métabolisme du cholestérol à l'intérieur de l'œil. L'approche qui n'a encore été testée que chez la souris — consisterait à cibler avec un collyre les cellules chargées d'évacuer le cholestérol. Celui-ci retournerait ainsi dans la circulation sanguine et ne s'accumulerait plus dans l'œil, empêchant la maladie de progresser. Cette idée inédite vient compléter l'arsenal des techniques en développement pour préserver ou restaurer la vision (lire Sciences et Avenir, n° 793, février 2013).
Rappelons que le cholestérol est un lipide impliqué dans de nombreux processus biochimiques. Mais quel rapport peut-il exister entre le cholestérol et le développement de vaisseaux sanguins de la rétine ? Il n'y a pas de lien entre DMLA et taux sanguin de cholestérol. La clef s'appelle en fait macrophages. Cette catégorie de cellules immunitaires joue en effet un rôle primordial dans l'élimination du cholestérol de l'œil. Or, les macrophages deviennent moins performants avec l'âge. Il en résulte des dépôts lipidiques qui entretiennent un foyer inflammatoire. Celui-ci favorise le développement de vaisseaux anormaux sous la rétine, responsables d'altérations sévères de la vision centrale. Pour l'heure, on ignore les mécanismes précis qui conduisent ces « vieux » macrophages à induire cette prolifération vasculaire. En revanche, de récents travaux ont montré qu'ils comptaient moins d'ABCAl, une protéine « transporteuse » permettant d'évacuer le cholestérol. Pour preuve, de « jeunes » macrophages dépourvus de ce même transporteur se comportent effectivement comme de « vieux » macrophages, favorisant une vascularisation anormale. D'où l'idée des chercheurs de « booster » l'expression d'ABCAl dans ces cellules afin qu'elles puissent se débarrasser de leur cholestérol et trouver la capacité à réguler la croissance vasculaire.
Pour ce faire, l'équipe de Rajendra Apte, du département d'ophtalmologie de l'École de médecine de l'université Washington à Saint-Louis (Missouri), a utilisé des gouttes ophtalmiques contenant un agoniste du LXR (Liver X Receptor). Cet agent stimule l'action d'un récepteur initialement découvert dans le foie et impliqué dans le métabolisme des lipides. Des études préalables ont en effet montré qu'il augmente la production du transporteur ABCA1 par les macrophages. Et les résultats des essais ont été encourageants : ainsi « dopées », les « vieilles » cellules se sont remises à fabriquer cet indispensable transporteur dans les mêmes proportions que les « jeunes ».
Restait à évaluer cette stratégie sur des souris chez lesquelles on a induit, sous l'effet d'un laser, une croissance anormale des vaisseaux sous-rétiniens, reproduisant ainsi ce que l'on observe chez les patients souffrant de DMLA. Là encore, les résultats ont été positifs. Administré en collyre, l'agoniste du LXR a permis de réduire significativement la prolifération vasculaire chez les souris traitées, les macrophages défaillants parvenant à nouveau à évacuer efficacement le cholestérol.
Il pourrait s'agir d'un progrès thérapeutique majeur
II faut désormais poursuivre ces recherches pour confirmer ces bons résultats, et surtout les appliquer à l'homme, ce qui prendra encore de longues années. S'ils se révélaient concluants, il s'agirait d'un progrès thérapeutique majeur dans la mesure où la fréquence de la DMLA augmente avec l'âge, représentant la principale cause de malvoyance chez les personnes de plus de 50 ans. Selon les chercheurs, « intervenir avant que la maladie ne soit à un stade avance avec des agents efficaces capables de stimuler la sortie du cholestérol de l'œil pourrait empêcher la progression de la maladie. Cette stratégie pourrait être déployée préventivement ».
Source : Marc Gozian Sciences et Avenir, n° 795, mai 2013, pp. 78-79.
12:29 Publié dans Biologie, Médecine | Tags : cellules souches, cellules souches embryonnaires, médecine régénératrice, rétine, thérapie | Lien permanent | Commentaires (3) | Facebook | | |
Élaboration d'une rétine en culture à partir de cellules souches embryonnaires
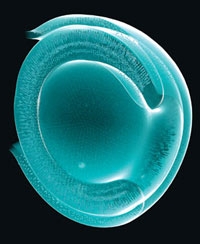 Élaboration d'une rétine en culture
Élaboration d'une rétine en culture
à partir de cellules souches embryonnaires
par Yoshiki Sasai[1]
Lors du développement embryonnaire, dans l'utérus, un petit amas de cellules identiques donne naissance aux divers types de cellules du corps humain. Elles s'arrangent en structures variées et finissent par former les organes. Un "programme" interne dirige cette progression depuis des éléments simples jusqu'à des tissus de forme et de taille appropriées.
Les biologistes aimeraient reproduire le développement embryonnaire en laboratoire, à la fois pour mieux comprendre ses mécanismes et pour élaborer des moyens le réparer et remplacer des tissus endommagés. Le succès est peut-être proche : de récents progrès laissent envisager la "culture d'organes" de remplacement à l'extérieur du corps ; ils pourraient être disponibles pour les opérations chirurgicales d'ici dix ans.
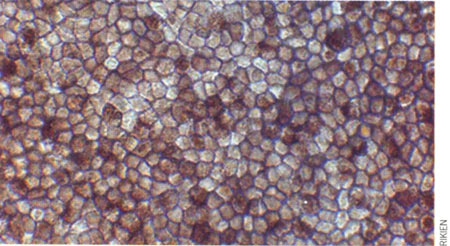
Cellules épithéliales pigmentées de la rétine
Mon optimisme se fonde sur des études sur les cellules souches – des divers types de cellules. Nous avons montré que des cultures de cellules souches peuvent donner naissance à une rétine ; cette structure clef de l'œil traduit la lumière en signaux électriques et chimiques, qui sont ensuite transmis au cerveau. Dans d'autres travaux, nous avons fait pousser du tissu cérébral (cortex) et une partie de l'hypophyse (une petite glande située à la base du cerveau). Pour ce faire, nous avons placé une mince couche de cellules dans une boîte de Pétri, où nous avons ajouté un cocktail de molécules présentes dans l'organisme. Ces cellules se sont développées en une structure tridimensionnelle. Nous espérons que du tissu rétinien produit de la sorte permettra de traiter plusieurs maladies de l'œil, telle la dégénérescence maculaire (la macula est la partie centrale de la rétine.
La formation de la rétine est un phénomène auto-organisé, fondé sur un programme interne des cellules.
À l'origine, nous cherchions à mieux comprendre la formation de la rétine. Nous savions qu'elle provient d'une partie du cerveau embryonnaire nommée diencéphale. Durant les premiers stades du développement embryonnaire, un segment du diencéphale s'étend pour former la vésicule optique, une structure en forme de ballon. Celle-ci s'invagine, formant la "cupule optique", qui deviendra la rétine (voir figure ci-dessous).
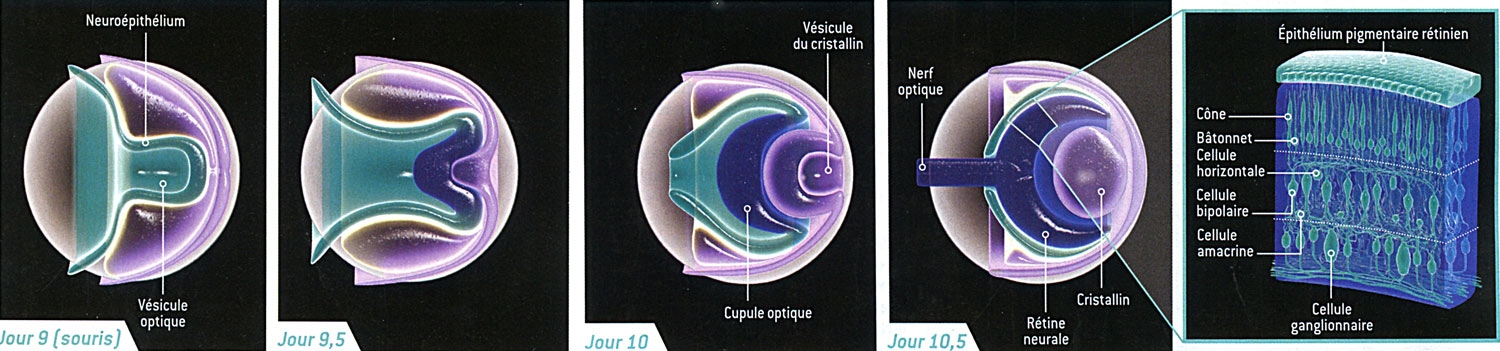
Naissance de la rétine chez l'embryon
L'œil se forme à partir d'un petit amas de cellules embryonnaires. Au début, une couche de tissu, le neuroépithélium, se replie vers l'extérieur, donnant naissance à la vésicule optique (jour 9). La paroi de ce bourgeon s'incurve ensuite vers l'intérieur (jour 9,5), et la vésicule du cristallin se développe (jour 10). Une partie de la vésicule optique engendre la cupule optique, à l'origine de la rétine et du nerf optique (jour 10,5). La rétine contient trois couches distinctes de cellules (à l'extrême droite), l'une d'elles étant constituée des photorécepteurs (les bâtonnets et les cônes). La rétine représentée ici, d'une taille de 250 micromètres est celle d'une souris. La rétine humaine est similaire, mais plus grosse, et sa formation est plus longue.
Depuis plus d'un siècle, les biologistes débattent sur le mécanisme de formation de la cupule optique. Une question importante concerne le rôle des structures voisines, telles que le cristallin et la cornée. Certains chercheurs pensent que le cristallin façonne la cupule optique en exerçant une pression sur la vésicule, tandis que d'autres affirment que la cupule peut se former sans lui et sans les autres tissus voisins.
Il est difficile d'observer les mécanismes physiologiques sous-tendant le développement sur des animaux vivants. Il y a environ dix ans, nous avons donc décidé de reproduire le développement de l'œil dans une boîte de Pétri. Pour ce faire, nous avons utilisé des cellules souches embryonnaires, que nous avons exposées à diverses substances chimiques connues pour être impliquées dans la formation de l'œil. Les cellules souches embryonnaires constituent le type le plus immature de cellules souches. Elles sont capables de se différencier en tous les types cellulaires du corps, des neurones jusqu'aux cellules musculaires.
Aucune technique existante ne permettait d'engendrer des organes à partir de cultures de cellules souches. Des essais avaient été réalisés en plaçant les cellules sur une sorte d'échafaudage artificiel, qui imitait la forme de l'organe ciblé (en l'occurrence une vessie ou un œsophage), mais ils n'avaient remporté qu'un succès mitigé. Nous avons donc tenté une approche différente.
En 2000, nous avons élaboré une méthode pour transformer des cellules souches embryonnaires de souris en différents types de neurones. Nous avons ensuite placé une monocouche de ces cellules souches dans une boîte de Pétri, avec des cellules dites nourricières : ces dernières transmettaient des signaux chimiques déclenchant la différenciation des cellules souches embryonnaires. Cette couche mince ne reproduisait pas les organes tridimensionnels réels, mais nous désirions déjà savoir si les molécules sélectionnées suffisaient à obtenir les types particuliers de neurones apparaissant précocement lors du développement de l'œil.
Après des débuts difficiles, nous avons développé une nouvelle méthode en 2005, qui nous a permis de dépasser le stade bidimensionnel : nous avons créé une suspension de cellules souches dans un milieu liquide, qualifiée de culture flottante. L'intérêt d'une telle culture tridimensionnelle est double. Tout d'abord, un agrégat de cellules est mieux à même de prendre la forme des tissus s'il est tridimensionnel que s'il est plat. Ensuite, la formation de structures complexes requiert de multiples communications entre cellules, et ces dernières interagissent plus facilement dans une culture tridimensionnelle.
Nous avons placé des cellules en suspension dans de petits réservoirs - comptant chacun en moyenne 3 000 cellules - au sein d'une boîte de Pétri. Initialement séparées les unes des autres, les cellules se sont liées avec leurs voisines. Nous avons ensuite déclenché la différenciation de ces petits agrégats de cellules en des progéniteurs neuronaux (des cellules neuronales immatures) présents à l'avant du cerveau. Les cellules ont alors commencé à échanger des signaux.
Trois à quatre jours plus tard, elles se sont organisées spontanément en une sphère creuse formée d'une couche cellulaire ; on parle de neuroépithélium. Nous avons nommé SFEBq cette méthode de culture (pour Serum free floating culture of embryoid body-like aggregate with quick reaggregation, littéralement culture sans sérum flottante d'agrégats embryonnaires se réagrégeant rapidement).
Dans l'embryon, les cellules neuroépithéliales forment des structures cérébrales spécifiques, après avoir reçu des signaux chimiques externes. L'un de ces signaux déclenche le développement du diencéphale, qui engendrera la rétine et l'hypothalamus (une aire cérébrale contrôlant l'appétit et de nombreuses fonctions physiologiques fondamentales). Après avoir fait s'agréger les cellules en sphères, nous avons provoqué leur différenciation en progéniteurs rétiniens - les précurseurs des cellules rétiniennes matures. Pour ce faire, nous avons ajouté à notre culture un mélange de protéines, incluant celles qui assurent ce rôle chez l'embryon.
Quelques jours plus tard, des structures semblables à la vésicule optique ont spontanément bourgeonné sur les sphères, avant de changer de forme : la structure finale ressemblait à la cupule optique de l'œil embryonnaire (voir figure ci-dessous). Comme la cupule optique des animaux vivants, cette structure issue de cellules souches embryonnaires se composait de deux parois (deux épithéliums), l'une extérieure et l'autre intérieure ; c'est cette dernière qui est à l'origine de la rétine.
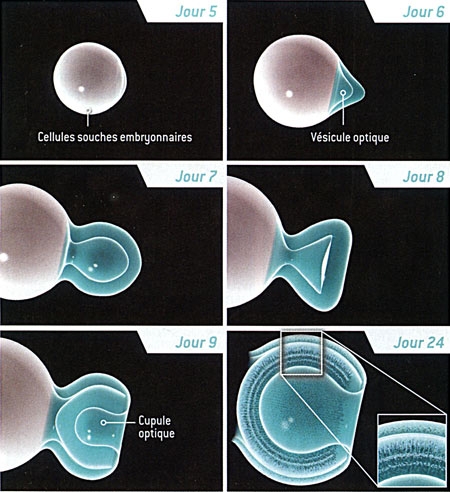
Formation de la rétine à partir d'une cellule souche
La formation d'une rétine à partir d'une culture de cellules souches embryonnaires ressemble à ce qui se produit dans l'utérus. Les cellules s'agrègent et commencent à former la vésicule optique "primitive" environ cinq jours après avoir été mélangées avec des substances nommées facteurs de croissance. Cette vésicule gonfle, puis forme la cupule optique. Le 24e jour, toutes les couches de la rétine sont visibles.
Ainsi, une structure ordonnée est née de la seule agrégation de cellules souches dans un récipient. Contrairement à ce qui se passe chez l'embryon, il ne se formait pas de cristallin ou de lentille au voisinage de la cupule optique. Nos travaux montrent donc qu'aucune force exercée par des tissus voisins (tel le cristallin) n'est nécessaire à la formation de la protorétine. Au moins in vitro, c'est un phénomène auto-organisé, fondé sur un programme interne des cellules. Ce programme est encodé dans leur ADN, et nous travaillons à préciser ses mécanismes d'action.
L'apparition spontanée d'une structure stratifiée
Le développement normal de la rétine, tel qu'on l'observe chez les embryons, s'est poursuivi dans nos cultures. En deux semaines supplémentaires, la cupule optique a grandi jusqu'à environ deux millimètres de diamètre, et l'épithélium monocouche interne est devenu une structure stratifiée, contenant les six catégories de cellules d'une rétine naturelle après la naissance. L'une des couches extérieures de cette structure était constituée de photorécepteurs et l'autre de cellules dites ganglionnaires, qui, dans le corps, relient la rétine au cerveau. Entre les deux se trouvaient plusieurs couches de cellules de liaison, nommées interneurones.
De nombreuses questions demeurent, notamment sur la façon dont une boule de cellules entre dans un programme de différenciation, qui aboutit à des tissus distincts. Tout au long du développement embryonnaire, des formes complexes apparaissent spontanément à partir d'amas de matière homogène - on parle de brisure de symétrie. En l'absence de ce phénomène, les divisions cellulaires de l'œuf fécondé n'engendreraient qu'une masse indifférenciée de cellules. Nos cultures de cellules souches embryonnaires qui s'auto-organisent devraient aider à mieux comprendre les mécanismes en jeu dans les brisures de symétrie.
On cherche également à mieux comprendre comment s'effectue le modelage de la cupule optique à partir d'une simple couche de cellules (l'épithélium rétinien). La force qui s'applique sur les cellules et la rigidité varient dans l'épithélium. En mesurant in vitro la direction de la force et la rigidité du tissu en différents endroits, nous avons découvert que la cupule optique se forme en plusieurs étapes. Tout d'abord, la rigidité de la protorétine diminue, ce qui accroît sa flexibilité. Dans le même temps, les cellules qui se trouvent à la jonction de l'épithélium (la boule dont a bourgeonné la rétine) et de la protorétine adoptent une forme en coin. Finalement, la protorétine commence à se replier vers l'intérieur en raison de sa rapide extension. Nous avons appliqué les conditions mécaniques correspondantes à une simulation sur ordinateur, et la forme de la cupule optique est apparue !
Pour y voir plus clair...
Nos travaux aideront-ils à lutter contre les troubles oculaires humains ? Nous avons progressé dans cette voie. En particulier, mon équipe a récemment recréé une cupule optique et un tissu neuronal à couches multiples à partir de cellules souches embryonnaires humaines. La même méthode de culture devrait être applicable à des cellules souches humaines dites pluripotentes induites - des cellules matures et différenciées dont on a « inversé » le développement pour qu'elles se comportent comme des cellules souches embryonnaires. Nous avons également inventé une méthode fiable de cryoconservation, qui permet de stocker du tissu rétinien issu de cellules embryonnaires humaines dans de l'azote liquide.
Ces travaux promettent des applications médicales. Par exemple, nous pourrions créer des rétines artificielles, qui aideront à étudier les maladies de l'œil et, peut-être, à développer des médicaments et des thérapies géniques pour inverser la dégénérescence rétinienne.
Trois catégories de dégénérescence rétinienne pourraient bénéficier de nos recherches : la dégénérescence maculaire, la rétinopathie pigmentaire et le glaucome. Ces maladies touchent des millions d'individus à travers le monde et altèrent des couches différentes de la rétine.
Dans la dégénérescence maculaire, un tissu nommé épithélium pigmentaire rétinien est endommagé : ce tissu tapisse le fond de la rétine et supporte les photorécepteurs, qui se détériorent également, en particulier dans la région centrale de la rétine.
Dans la rétinopathie pigmentaire, les bâtonnets (un type de photorécepteurs) disparaissent progressivement pendant plusieurs années. La maladie se manifeste d'abord par une cécité nocturne, ou héméralopie, puis le champ visuel se réduit jusqu'à se limiter à une petite zone au centre, avant que le patient ne devienne totalement aveugle.
Enfin, le glaucome endommage les cellules ganglionnaires, qui relient la rétine au cerveau via le nerf optique.
De ces trois maladies, la dégénérescence maculaire semble la plus susceptible d'être traitée par une thérapie cellulaire. Pour obtenir les cellules nécessaires au remplacement de l'épithélium pigmentaire rétinien endommagé, on transforme des cellules souches embryonnaires humaines ou des cellules souches pluripotentes induites, au sein de cultures conventionnelles ou flottantes. Des essais cliniques à petite échelle ont débuté aux États-Unis et sont prévus dans d'autres pays : les cellules d'épithélium pigmentaire issues de cellules souches sont injectées sous la rétine avec une fine aiguille, afin de remplacer au moins une partie du tissu endommagé.
Pour la rétinopathie pigmentaire, la thérapie cellulaire est une perspective plus lointaine. À l'inverse d'une culture classique, notre méthode peut engendrer une couche dense de bâtonnets facilement transplantable. En revanche, ces photorécepteurs doivent s'intégrer dans le circuit neuronal de l'œil - ce qui n'était pas nécessaire pour l'épithélium, simple tissu de soutien. Ainsi, ils doivent se connecter à un autre type de cellules sensorielles, les cellules bipolaires, et nous ne savons pas encore comment réaliser cette connexion.
Le glaucome pourrait être la plus difficile des trois maladies à traiter par thérapie cellulaire. Des cultures de cellules souches embryonnaires peuvent produire les cellules ganglionnaires nécessaires, mais dans l'œil postnatal, le nerf optique ne se reforme pas spontanément. Autrement dit, une fois les cellules ganglionnaires transplantées, on ne sait pas connecter leurs axones (les prolongements des neurones qui forment le nerf optique) aux cellules cérébrales.
Notre méthode a décuplé les possibilités des tissus formés à partir de cellules souches embryonnaires. Reste à découvrir ce qu'elle peut nous apprendre sur les mécanismes qui conduisent d'une cellule unique à un organe aussi complexe que l'œil...
BIBLIOGRAPHIE
Y. Sasai (2013). – Une rétine en culture. Pour la Science, n° 426 avril 1993, pp. 58-63.
T. Nakano et al. (2012). - Self-formation of optic cups and storable stratified neural retina from human ESCs, Cells Stem Cell, vol. 10, pp. 771-785.
D Schwartz et al. (2012). - Embryonic stem cell trials for macular degeneration : a preliminary report, Lancet, vol. 379, pp. 713-720, 2012.
M. Eiraku et al. (2011). - Self-organizing optic-cup morphogenesis in three-dimensional culture, Nature, vol. 472, pp. 51-56.
Les alternatives thérapeutiques aux cellules souches
par Serge Picaud et José Sahel, Institut de la vision, Paris
Si les cellules souches ouvrent de nouvelles perspectives thérapeutiques pour les patients aveugles, une autre technique en est déjà au stade commercial : celle des prothèses rétiniennes. Ces dispositifs visent à stimuler artificiellement une rétine ayant perdu ses photorécepteurs. Les images, filmées par une caméra fixée sur une paire de lunettes, sont d'abord transformées en matrices d'impulsions électriques. Celles-ci sont ensuite transmises à des électrodes implantées sur la rétine résiduelle (voir la figure ci-dessous).
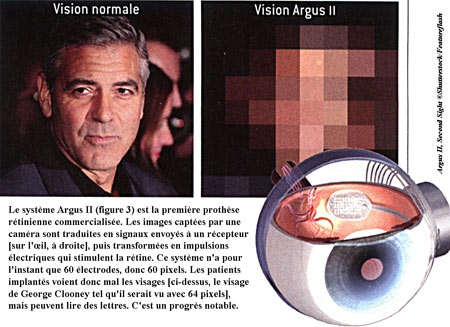
Commercialisée depuis deux ans en Europe, la première prothèse rétinienne opérationnelle a reçu l'autorisation de mise sur le marché aux États-Unis en février 2013. En France, quatre patients aveugles ont participé aux essais cliniques. Cependant, les images perçues ne sont constituées que de 60 points ou pixels, puisque l'implant n'a que 60 électrodes : c'est insuffisant pour bien voir une image complexe, tel un visage, mais suffisant pour une image simple, de sorte que certains patients sont capables de lire des lettres d'environ 10 centimètres de hauteur.
De nombreux chercheurs tentent d'augmenter le nombre de pixels, chaque électrode devant activer une zone disjointe de sa voisine. Ainsi, à l'Institut de la vision, on poursuit plusieurs objectifs : le développement de nouvelles formes tridimensionnelles pour les implants, avec l'ESIEE (École supérieure d'ingénieurs en électrotechnique et électronique) et le CEA; l'utilisation de matériaux semi-conducteurs différents, tels le graphène et le diamant (projet européen NEUROCARE) ; l'amélioration du codage des informations visuelles.
Une piste alternative, la thérapie optogénétique, est inspirée d'algues unicellulaires et de bactéries dotées de protéines photosensibles. Ces protéines sont des canaux ioniques dont l'ouverture est contrôlée par la lumière. L'idée est d'insérer le gène qui les code dans les neurones non photosensibles de la rétine, qui pourraient alors être activés par la lumière.Ainsi, chaque cellule produisant les protéines jouerait le rôle d'un pixel. Ces protéines étant moins performantes que nos pigments visuels, les aveugles devront porter des lunettes rehaussant la luminance.
Ainsi, dans le cadre du projet européen RETICIRC, nous avons rendu la vue à des souris aveugles grâce à une protéine bactérienne. Nous avons ensuite montré que cette protéine permet l'activation de cellules rétiniennes humaines par la lumière, grâce à des cultures de cellules prélevées post mortem. Avant de passer à une application clinique, nous devrons vérifier que la thérapie génique visant à faire exprimer la protéine bactérienne n'entraîne pas d'inflammation ou de rejet chez le primate.
[1] Yoshiki SASAI est directeur du groupe d'organogenèse et de neurogenèse au Centre de biologie du développement du RIKEN, à Kobe, au Japon. Plus d'information sur Y. Sasai et ses recherches.
12:05 Publié dans Biologie, Médecine | Tags : cellules souches embryonnaires, rétine, culture organotypique, macula, dmla, rétinopathie pigmentaire, glaucome, prothèse rétinienne | Lien permanent | Commentaires (2) | Facebook | | |
21/07/2012
Cellules souches : obtenir des neurones à partir de fibroblastes
Obtenir des neurones
à partir de fibroblastes
Les cellules de l'organisme, y compris les neurones possèdent la même information génétique, mais elle n'est pas lue de la même façon selon le type cellulaire. On sait aujourd'hui agir sur cette lecture, notamment à l'aide de facteurs de transcription, des protéines qui se fixent sur l'ADN et régulent sa transcription en ARN. Grâce à un seul de ces facteurs, l'équipe de Karen Ring et Yadong Huan, de l'Université de Californie (San Francisco) a réussi à obtenir des réseaux de neurones à partir de fibroblastes humains et de souris.
Pour transformer les fibroblastes en d'autres cellules, on les place dans un milieu de culture adapté, qui recrée l'environnement physico-chimique des cellules qu'on cherche à obtenir, et l'on provoque la sur-expression des facteurs de transcription responsables de la différenciation visée.
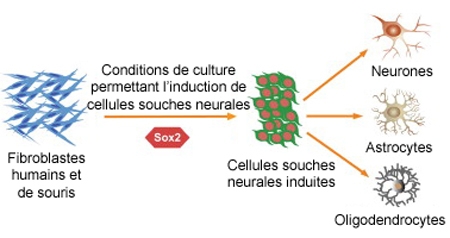
Protocole de l'expérimentation
Cette dernière opération consiste à faire produire en masse ces facteurs de transcription par la cellule. L'équipe de Y. Huang et de Karen Ring n'en a utilisé qu'un seul, nommé Sox2. D'autres équipes avaient également obtenu des neurones, mais elles avaient fait appel à un nombre bien plus important de facteurs de transcription.
En quelques jours, les fibroblastes se sont transformés en cellules souches neurales induites (iNSCs). Celles-ci se sont divisées et se sont différenciées en astrocytes, oligodendrocytes et en neurones fonctionnels. En moins d'un mois, ces neurones se sont associés en réseaux.
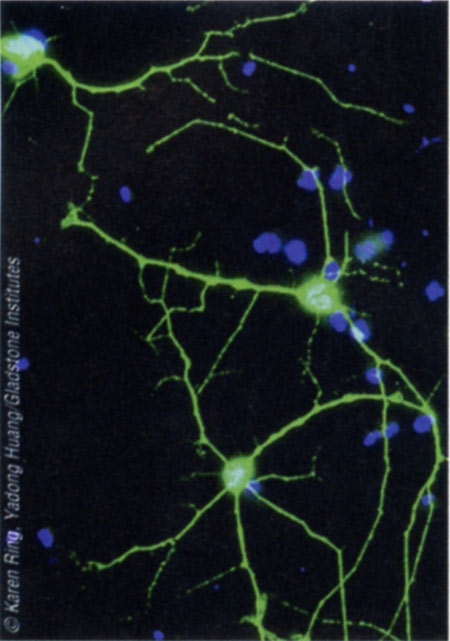
Neurones issus de la transformation de fibroblastes
Pour vérifier que les cellules reprogrammées ne risquaient pas de dégénérer en cellules cancéreuses, l'équipe américiane les a greffées dans le cerveau d'une souris. Elles s'y sont intégrées sans causer de tumeur. Ainsi, par ce type de reprogrammation, les iNSCs auto-renouvelables et multipotentes sans potentiel tumorigène peuvent être générées directement à partir de fibroblastes. Ces cellules seront peut-être utiles dans la lutte contre les maladies neurodégénératives, telles celles d'Alzheimer et de Parkinson ; elles permettront de tester les médicaments élaborés.
Source :
K. King et al. (2012). - Direct Reprogramming of Mouse and Human Fibroblasts into Multipotent Neural Stem Cells with a Single Factor. Cell Stem Cell, 7 juin 2012.
21:47 Publié dans Actualité des Sciences, Biologie, Médecine | Tags : cellules souches, fibroblastes, neurones, astrocytes, oligodendrocytes, cellules souches neurales induites, inscs | Lien permanent | Commentaires (0) | Facebook | | |
18/06/2012
OGM : la France maintient le bon cap
 OGM : la France maintient le bon cap
OGM : la France maintient le bon cap
France Nature Environnement (FNE) salue la décision de la France qui, lors du Conseil européen de l’environnement du 11 juin 2012, a refusé que soient modifiées les modalités d’autorisation de mise en culture d’OGM. Bien au-delà de ce constat, il importe pour FNE que soient rappelés enjeux de société et stratégies.
Une nouvelle fois, il s’est agit de déjouer un marché de dupes consistant à accorder aux États membres la possibilité d’interdire sur leurs territoires des cultures d’OGM, pour faciliter les autorisations de mise en culture au niveau communautaire par un allègement des procédures de leur évaluation !
FNE a dénoncé a plusieurs reprises les carences de ces procédures menées par l’Agence européenne de sécurité alimentaire (AESA / EFSA), ainsi que son fonctionnement, alors que ses avis conditionnent les autorisations accordées par la Commission européenne (CE). Rappelons que, en France, le Comité scientifique du Haut Conseil des Biotechnologies et l’ANSES ont contesté la validité de ces expertises européennes, notamment en raison de l’insuffisance de la puissance statistique des tests présentés par la firme MONSANTO au sujet de son maïs MON 810 en question (tests examinés par l'AESA).
Celle-ci reproche à la France de ne pas apporter de preuves scientifiques nouvelles dans son dossier d’interdiction du MON 810 renouvelé en février 2012, alors que l’une de ces preuves est apportée par son évaluation défaillante !

Selon Lylian Le Goff, pilote de la mission Biotechnologies de FNE : « Des procédures d’évaluation moins contraignantes, reposant sur le dogme aberrant de l’équivalence en substance entre variété OGM et non OGM, faciliteraient autorisations et cultures dans les quelques états favorables aux OGM et, par conséquent, la contamination des Etats soucieux de préserver l’intégrité de leur biodiversité et la spécificité de leurs territoires, notamment par la libre circulation des semences défendue par l’OMC. Voilà pourquoi il est vain de permettre à un état d’interdire les OGM sur son territoire sans un renforcement de l’évaluation de leurs risques toxicologiques mais aussi de leurs impacts socio-économiques».
Il importe effectivement de maintenir le cap et que l’avis unanime émis en décembre 2008 par le Conseil européen des ministres de l’environnement soit enfin appliqué : «renforcer et harmoniser les méthodes d’évaluation des risques des OGM en Europe ».
18:59 Publié dans Biologie, Environnement-Écologie, Nature et santé, Pollution | Tags : fne, france nature environnement, monsanto, mon810, aesa, efsa, commission européenne | Lien permanent | Commentaires (0) | Facebook | | |
20/05/2012
Médecine et génomique à l'Institut Pasteur
 Médecine et génomique à l'Institut Pasteur
Médecine et génomique à l'Institut Pasteur
Nous ne sommes pas tous égaux devant les maladies
"Tous les hommes sont égaux, mais certains sont plus égaux que d'autres" écrivait Georges Orwell. Ceci est un peu vrai face à la maladie. Si certaines pathologies sont provoquées par des mutations dans un gène unique, la plupart des maladies courantes ont à la fois une composante environnementale (nutrition, activité physique, etc.) et génétique – en témoignent les questions de nos médecins sur nos antécédents familiaux. Et qu'il s'agisse de diabète, d'asthme, de maladie d'Alzheimer, de cancers ou de maladies infectieuses, de multiples variations dans de multiples gènes interviennent.

Des facteurs génétiques de prédisposition ou de protection
Certaines nous rendent plus vulnérables face à une maladie donnée – elles touchent des gènes "de prédisposition" – d'autres au contraire nous en protègent. Une mutation dans les gènes BRCA1 ou BRCA2 confère par exemple aux femmes qui la portent (2 sur 1 000) un risque de 40 à 80 % selon l'âge de développer un cancer du sein. Les rares individus porteurs d'une mutation du gène CCR5 sont eux résistants à l'infection par le virus du sida. D'autres gènes conditionnent la réponse aux traitements. Ainsi, 11 % des patients réagissent mal à une molécule, la mercaptopurine, utilisée pour traiter certaines leucémies. Chez eux, une activité réduite de l'enzyme TMPT freine l'élimination du médicament, induisant des effets très toxiques. Un test génétique est donc désormais recommandé aux États-Unis afin d'éviter ce traitement ou de rappliquer à très faibles doses chez ces patients.
Pour appréhender l'ampleur des recherches sur ces gènes de sensibilité aux maladies ou aux traitements et les perspectives médicales qui en découlent, il faut revenir sur la fantastique épopée scientifique qui a marqué le début du XXIe siècle.

Naissance de la génomique
Tout a commencé par l'aboutissement d'un projet titanesque, qui dura 13 ans : le Projet Génome Humain. Achevé en 2003, il révélait l'intégralité du texte d'un ADN humain – sa "séquence" – soit 3 milliards de lettres[1].
Il ouvrait l'ère de la génomique, rendant possible l'exploration des quelque 20 500 gènes contenus dans le génome humain, facilitée d'année en année par l'automatisation des techniques de séquençage et les progrès de la bio-informatique. Il a montré qu'il y a très peu de différences génétiques entre les individus ou les populations et pas de différences majeures entre groupes ethniques, abolissant définitivement le concept de race. Les génomes de deux individus pris au hasard dans la population mondiale ont en moyenne à peine 0,1 % de différences. Mais sur les trois milliards de lettres du texte de l'ADN, ce 0,1 % correspond tout de même à 3 millions de variations... dont bon nombre expliquent nos différences de vulnérabilité face aux maladies.
Nos différences génétiques cartographiées
Comment les étudier ? Dans la foulée du projet Génome Humain, un autre grand programme international – Hap-Map –, a dressé un catalogue des différences et des similarités génétiques entre individus. À partir des génomes de 270 individus d'origines africaine, asiatique et européenne, les variations génétiques les plus fréquentes (des changements ponctuels d'une lettre dans le texte de l'ADN, présents chez plus de 5 % des individus) ont été répertoriées. Trois millions de telles variations ont été cartographiées. Ces "balises" dans l'ADN ont permis de vastes études d'association sur des génomes entiers, portant chacune sur des centaines voire des milliers d'individus : les GWAS (genome-wide association studies). Schématiquement, un panel d'individus sains est comparé à un panel d'individus touchés par une maladie donnée, sur la base du catalogue Hap-Map : si certaines variations génomiques sont beaucoup plus fréquentes chez les malades, elles sont associées à la pathologie. En janvier 2012, plus de 1000 "GWAS" avaient été publiées, mettant en évidence plusieurs centaines de nouveaux facteurs génétiques de risque pour des maladies courantes : diabète, maladies cardio-vasculaires, maladies neuro-dégénératives (Alzheimer, Parkinson), cancers, maladies mentales comme la schizophrénie ou l'autisme (voir encadré), maladies infectieuses (hépatites B, C, sida, tuberculose…).

Le projet "1000 génomes"
Ainsi, plus d'axes de recherche totalement inattendus et de pistes thérapeutiques nouvelles ont été identifiés depuis l'an 2000 qu'au cours du XXe siècle, et révolution de la génomique ne cesse d'apporter des outils toujours plus précis pour les recherches sur notre vulnérabilité aux maladies. Le projet "1000 génomes" lancé en 2008 vise désormais à fournir un grand nombre de séquences complètes de génomes humains à partir de toutes les populations du globe. Mais aussi à répertorier dans ces génomes des millions d'autres variations, dites "rares" (présentes chez moins de 0,5 % de la population), et divers "accidents" dans l'ADN qui n'ont pas à ce jour été cartographiés, comme de petites portions manquantes (délétions) ou surnuméraires (insertions). "Les chercheurs qui ont trouvé des régions du génome associées à une maladie pourront regarder ces données et avoir un panorama presque complet des variations génétiques dans ces régions pouvant contribuer directement à la maladie", souligne un des auteurs du projet. À ce jour, 2 500 échantillons d'ADN provenant de 27 populations à travers le monde doivent être analysés.
Vers une médecine prédictive et personnalisée ?
Côté applications, la révolution médicale annoncée au début des années 2000, qui permettrait de prédire à chacun quelles maladies il risque de développer et de proposer des traitements à la carte en fonction du profil génétique, n'en est qu'à ses prémices. Mais pour beaucoup, il ne s'agit plus de savoir si elle surviendra, mais quand. Un des freins tient au fait que l'on s'attendait à découvrir quelques gènes majeurs associés aux maladies, et qu'on en a trouvé des dizaines, voire des centaines : en août dernier, une équipe internationale annonçait avoir découvert 29 nouveaux variants génétiques associés à la sclérose en plaques, s'ajoutant à 23 variants déjà connus... soit une cinquantaine identifiés pour cette seule maladie ! Il reste par ailleurs difficile d'évaluer parmi les gènes de "prédisposition" à une maladie lesquels sont les plus "prédictifs" d'un risque. Aussi, les sociétés qui proposent aujourd'hui sur Internet des tests génétiques révélant nos risques de maladies à partir d'un échantillon de salive sont-elles montrées du doigt pour leur manque de fiabilité... Au fil des avancées, une médecine prédictive plus fiable verra cependant le jour avec à la clé des mesures de prévention mieux ciblées. L'avancée des connaissances en pharmacogénomique, l'étude des facteurs génétiques individuels en jeu dans la réponse aux traitements, en plein développement, conduira parallèlement à l'essor de la médecine personnalisée : déjà, plusieurs traitements existent accompagnés de tests génétiques "compagnons", qui conditionnent leur utilisation chez un patient selon qu'il est "bon répondeur" ou bien à risque d'effets indésirables.

1 000 dollars pour séquencer un génome
Un élément de taille est venu récemment conforter cette tendance à envisager une médecine du futur tenant compte du profil génétique du patient : la chute drastique des coûts du séquençage. Si en 2001 des centaines de millions de dollars étaient nécessaires pour obtenir la séquence génomique complète d'un individu, ce prix tombait à 50 000 dollars en 2009. En février 2011, une société américaine annonçait dans la revue scientifique Nature réduire ce coût à 1000 dollars. Le séquençage d'un génome serait bientôt à un prix si abordable qu'il pourrait être généralisé à tout un chacun. À quand notre séquence génomique dans notre carte Vitale ? Les questions éthiques et sociétales soulevées par cette perspective – la protection des données génétiques vis-à-vis des assurances ou des employeurs par exemple – deviennent urgentes à résoudre...
En attendant, chaque variation génétique associée à une maladie est une piste nouvelle pour les chercheurs. Il faut alors comprendre le mécanisme biologique dans lequel le gène impliqué intervient, trouver le moyen d'agir en conséquence pour traiter la maladie... La décennie précédente a fourni une avalanche de données génétiques associées aux maladies : seul le sommet de cet iceberg d'informations a été exploré. Son exploitation fournira sans doute bien des solutions inédites face aux maladies qui nous menacent.
Génomique et paludisme : vers de nouvelles pistes de recherche
Près d'un million de morts et 500 millions de cas par an, des traitements de moins en moins efficaces : le paludisme, un des premiers fléaux sanitaires de la planète, est une véritable urgence pour la recherche et l'étude des facteurs de sensibilité à cette infection apporte l'espoir de nouvelles voies d'investigation.
À l'Institut Pasteur, l'unité de Génétique fonctionnelle des maladies infectieuses, dirigée par Anavaj Sakuntabhai, avait déjà contribué à la découverte d'une mutation génétique conférant une résistance au paludisme chez 18 à 25 % des individus en Asie du Sud-Est, après une étude sur 3 500 personnes en Thaïlande. "Cette mutation est située dans un gène impliqué dans le stress oxydatif : ce phénomène intervient dans le vieillissement et la destruction des globules rouges, lieux de multiplication des parasites", explique Jean-François Bureau, un des chercheurs de l'unité.
Aujourd'hui, l'équipe pasteurienne lance une vaste étude de recherche de gènes associés à l'infection (GWAS[2]). Sa source : les échantillons d'ADN de plus d'un millier d'habitants des villages de Dieimo et de Ndiop au Sénégal – suivis depuis 1990 par l'Institut Pasteur de Dakar et l'Institut de Recherche pour le Développement (IRD) –, et de 600 personnes au Pérou. "Nous allons cibler nos recherches en définissant des sous-groupes dans ces populations exposées au paludisme : en fonction du nombre d'accès palustres, de leur sévérité, de l'âge – tes enfants étant les plus touchés –, etc." précise Richard Paul, autre chercheur de l'équipe. "Grâce au suivi de ces habitants depuis 20 ans, nous disposons de quantité de données, parfois depuis la naissance. À chaque fois que nous découvrirons une mutation dans un gène, nous saurons ainsi à quel type de cas clinique elle est associée." Les premiers résultats devraient être disponibles dans un an.
Autisme : la part des gènes
Dès les années 70, des études chez les jumeaux mettaient en évidence une composante génétique dans l'autisme, trouble envahissant du développement qui touche un enfant sur 100. En 2003, les premiers gènes associés à des cas d'autisme et de syndrome d'Asperger (autisme avec langage et sans retard mental) étaient découverts à l'Institut Pasteur dans l'unité de Génétique humaine et Fonctions cognitives dirigée par Thomas Bourgeron. Aujourd'hui, plus de cent gènes sont associés à la maladie. Quelle part jouent-ils ?
"C'est très complexe", explique Thomas Bourgeron. "D'abord, il n'y a pas "un" autisme, mais un spectre de troubles autistiques, donc quantité de patients différents et autant de cas particuliers. Ensuite, l'impact de chaque gène diffère. Par exemple, la présence de mutations dans le gène SHANK3 que nous avons découvert signe quasiment toujours un autisme.
Ces cas-là sont "très génétiques". Chez d'autres patients, les mutations identifiées sur d'autres gènes n'expliquent pas à elles seules la maladie, qui relève probablement d'autres facteurs génétiques non identifiés et peut-être aussi de facteurs épigénétiques et environnementaux. Quoiqu'il en soit, ces gènes de vulnérabilité sont autant de pistes de recherche pour mieux comprendre les autismes." Dans le laboratoire de l'Institut Pasteur, généticiens côtoient neurobiologistes et psychiatres. Il ne s'agit pas seulement d'isoler des gènes associés à l'autisme mais de comprendre à quoi ils servent. "Pour définir la fonction de gènes récemment Impliqués, nous cultivons actuellement des neurones avec des cellules exprimant ces gènes mutés" explique Thomas Bourgeron. "Nos résultats préliminaires montrent des différences dans la taille des neurites, ces prolongements neuronaux pouvant donner de nouvelles connexions".
D'autres gènes impliqués interviennent dans le fonctionnement des points de communication entre les neurones, les synapses. Des images en 3D de cerveaux d'enfants autistes, reconstituées à partir d'IRM, sont aussi analysées en vue de relier d'éventuelles modifications aux mutations génétiques trouvées chez ces patients. C'est donc une cascade d'études qui découle des découvertes de gènes de vulnérabilité, avec à la clé l'espoir d'améliorer la prise en charge des enfants touchés.
Source : La lettre de l'Institut Pasteur, mai 2012.
15:04 Publié dans Biologie, Médecine | Tags : médecine, génomique | Lien permanent | Commentaires (0) | Facebook | | |
09/03/2012
Régénération de la rétine par cellules souches embryonnaires
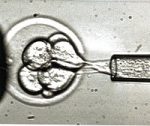 Régénération de la rétine humaine
Régénération de la rétine humaine
par cellules souches embryonnaires
(mise à jour du 25/01/2015)
Dans leurs livraisons de mars 2012, les revues Sciences & Vie (n° 1134) et Sciences et Avenir (n° 781) se sont fait l'écho d'une avancée thérapeutique fondamentale : pour la première fois, une thérapie a été mise au point à partir de cellules souches humaines : les rétines de deux patients ont été traitées par des cellules souches embryonnaires humaines (CSEh) par une équipe américaine. Encadrée par l'agence sanitaire américaine, l'opération réalisée le 12 juillet 2011 à l'Institut ophtalmologique Jules-Stein de Los Angeles dépasse de très loin la simple réparation de deux rétines et concerne la régénération des organes humains.
Ce sont les premiers blastomères issus de la segmentation des embryons surnuméraires après 5 ou 6 jours de développement non utilisés par les couples pour une fécondation in vitro. Contrairement aux cellules souches que l'on trouve dans certains organes adultes qui ont un pouvoir de différenciation limité, les cellules souches embryonnaires(CSEh) sont des cellules pluripotentes car elles sont à l'origine de toutes les cellules de l'organisme (voir dans ce même blog l'article : Cellules souches et rajeunissement).
Ces cellules pluripotentes possèdent le plus enviable de tous les pouvoirs, celui de générer la totalité des organes humains. Cerveau, muscles, cœur, poumons… Rien ne leur est impossible. Depuis leur découverte en 1998 à l'intérieur d'embryons de cinq ou six jours, les cellules souches embryonnaires (CSEh) ont fait l'objet de vastes programmes de recherches. Mais elles n'étaient pas encore sorties des laboratoires malgré quelques effets d'annonce jamais vérifiés.
L'opération américaine de de la rétine
Sous la rétine (1), 50 000 cellules pigmentaires (obtenues à partir de cellules souches embryonnaires (CSEh)) ont été injectées. Seule une petite zone a été traitée pour limiter les risques en cas d'effets secondaires, l'imagerie médicale permet d'observer, en coupe, le fond de la rétine (2). La petite surépaisseur visible avec un fort grossissement (3), correspond au point d'injection et témoigne que les cellules ont bien survécu et colonisé la zone. Même si la surface réparée est peu étendue, les patients estiment que leur vision s'est améliorée.
D'après Jean-Pierre Hubschman, un ophtalmologiste d'origine française de l'équipe, la manipulation était très simple : "Il s'agissait de légèrement soulever la rétine du patient, puis d'injecter quelques gouttes de liquide ". Mais ce liquide contient plus de 50 000 cellules pigmentaires, indispensables à la bonne réception de la lumière dans l'œil.
Ces cellules rétiniennes ont été obtenues grâce à la transformation d'environ 2 000 cellules souches embryonnaires (CSEh) mises en culture et différenciée par une méthode mise au point par la firme américaine de biotechnologie Advanced Cell Technology (ACT). Ainsi, injectées en un point précis, les cellules cultivées ont pu coloniser et retapisser, en partie du moins, le fond de la rétine des deux patients, une septuagénaire atteinte par une forme de dégénérescence maculaire liée à l'âge (DMLA), et un homme touché par la maladie de Stargardt, une dégénérescence maculaire héréditaire.
Quelques mois plus tard, les techniques d'imagerie de la rétine montrent que les cellules injectées sont toujours visibles dans la zone traitée et les deux patients estiment avoir bénéficié d'une amélioration de la vue, même s'ils continuent de souffrir de formes très avancées de leur maladie. Jean-Pierre Hubschman reste particulièrement prudent : "L'immense espoir suscité par l'annonce d'une thérapie contre ces deux maladies incurables ne doit pas faire croire au miracle. Avec seulement deux patients opérés, c'est encore trop tôt pour affirmer que nous avons réussi. Même si les résultats semblent très encourageants." Depuis, d'ailleurs, d'autres malades ont été opérés aux États-Unis et en Angleterre. La totalité des résultats est attendue pour 2014. Et, dans le même temps, d'autres essais sur des organes aussi divers que le cœur, le cerveau ou le pancréas pourraient très vite débuter. Cette fois, pas de doute, l'aventure des cellules souches embryonnaires est bien partie.
Une crainte subsiste car le pouvoir de différenciation des cellules souches embryonnaires (CSEh) a un revers : on les suspecte de pouvoir engendrer des tumeurs complexes à croissance rapide, les tératomes. Et c'est pour limiter ce risque qu'ACT a préféré miser sur une implantation dans l'œil, un organe non vital. Un choix d'autant plus judicieux que les réactions de rejets immunitaires y sont également très limitées. Les deux opérations pionnières de la rétine se devaient donc déjà de démontrer, avant même l'efficacité de la thérapie, l'innocuité des cellules souches embryonnaires.
"C'est une annonce très enthousiasmante", confirme Marc Peschanski, de l'unité I-Stem de l'Inserm au Génopole d'Évry (Essonne), l'un des 21 centres autorisés en France à mener des recherches sur ces cellules, "l'objectif intitial de l'étude a déjà été atteint. Six mois après l'opération, les cellules greffées semblent s'être parfaitement intégrées dans la rétine".
La DMLA« sèche» dans le viseur
Par ailleurs, la piste de la greffe de cellules rétiniennes dérivées de cellules souches embryonnaires humaines semble se confirmer (Sciences et Avenir, n° 816, février 2015, p. 96). Publiés en octobre dernier dans The Lancet, les derniers résultats d'une étude américaine dirigée par le Pr. Robert Lanza de la firme Advanced Cell Technology, montrent en effet une amélioration globale de la vision des patients greffés atteints de DMLA, et ce sans apparition d'effets indésirables irréversibles au bout de 22 mois.
Dans le même article, le magazine pose la question : Le Lampalizumab : un médicament efficace ? Capables de réduire de 20 % la progression des lésions causées par la forme « sèche » et avancée de la dégénérescence maculaire liée à l'âge (DMLA) — plus courante que sa forme « humide » et jusqu'ici sans traitement —, les injections de Lampalizumab « constituent un nouvel espoir », soutient le Dr Jean-François Girmens, ophtalmologiste à l'hôpital des Quinze-Vingts à Paris. Tout juste entré en phase III de tests dans 24 pays, cet inhibiteur d'une partie du complément (ensemble de protéines agissant sur notre réponse immunitaire) ne devrait toutefois pas être vendu avant 2017, prévient le laboratoire Roche.
La voie japonaise avec l'uilisation de cellules iPS (cellules souches adultes)
Devenues célèbres en octobre 2012 grâce au prix Nobel de médecine décerne à leur découvreur, le Pr Shinya Yamanaka, les cellules souches pluripotentes induites (ou cellules iPS) occuperont à nouveau le haut de l'affiche en 2013. Des chercheurs japonais du centre Riken lanceront en effet le premier essai thérapeutique sur l'homme faisant appel à ces cellules dans le traitement de la DMLA.
Les iPS se positionnent ainsi comme une alternative concrète aux cellules souches embryonnaires humaines (CSEh), celles-là mêmes qui laissent entrevoir depuis des années l'avènement d'une médecine régénératrice, mais dont l'utilisation se heurte aux problèmes éthiques posés par la question du statut de l'embryon. Les iPS peuvent être vues comme de « fausses » cellules embryonnaires : elles auraient les mêmes capacités de multiplication et de différenciation, mais sans provenir d'embryons. Car il s'agit de cellules différenciées adultes - par exemple de banales cellules de peau - « rajeunies » jusqu'au stade embryonnaire sous l'action d'un cocktail de jouvence mis au point par Shinya Yamanaka. Outre le traitement de la DMLA, le Japonais fonde de grands espoirs sur l'utilisation des iPS pour réparer les lésions de la moelle épinière.
L'espoir d'une thérapie
De nombreux organes sont potentiellement concernés par le pouvoir réparateur des cellules souches embryonnaires. Ainsi, Michel Pucé (Inserm) pourrait bien être le premier à se lancer dans des essais sur l'insuffisance cardiaque. "Les protocoles sont en phase de finalisation, explique le chercheur. Nous attendons le feu vert des agences sanitaires." Ces essais sur le cœur sont d'autant plus attendus qu'aucun résultat n'a pu encore être obtenu jusqu'à présent lors des nombreux essais de thérapies cellulaires.
Autre projet en cours : des tests sur des cellules pancréatiques pour traiter le diabète. Reste que, en dehors des essais menés sur la rétine, un seul autre projet a été homologué dans le monde: le laboratoire américain Neuralstem s'intéresse au potentiel réparateur des cellules souches contre la sclérose latérale amyotrophique, une maladie neurovégétative. Et les premières injections ont eu lieu. Quoiqu'il en soit, l'absence d'effets secondaires officiellement constatés dans l'essai d'ACT devrait rassurer les agences sanitaires encore prudentes.
Sources :
Tourbe C. (2012). - Cellules souches embryonnaires Sciences & Vie, mars 2012 n° 1134, pp.68-72.
Kaldy P. (2012). - Les cellules souches peuvent rendre la vue, Sciences et Avenir, mars 2012, n° 781, p. 34.
10:28 Publié dans Biologie, Médecine, Sexualité-embryologie | Tags : cellules souches, cellules souches embryonnaires, médecine régénératrice, moelle osseuse, greffe d'organe, rétine, thérapie | Lien permanent | Commentaires (6) | Facebook | | |
10/05/2011
OGM : de la toxine Bt retrouvée dans le sang de femmes enceintes et de leurs fœtus au Québec
OGM : de la toxine Bt retrouvée dans le sang de femmes enceintes et de leurs fœtus au Québec

D'après Greenpeace, une étude indépendante réalisée à Sherbrooke auprès de femmes enceintes, de leurs fœtus et de femmes non enceintes révèle la présence de résidus de pesticides issus d’aliments transgéniques dans le sang de chacun de ces groupes. Des résidus de glyphosate et de glufosinate (herbicides couramment utilisés lors de la culture de certaines plantes génétiquement modifiées) ont également été retrouvés.
Une étude scientifique publiée en avril par des chercheurs de l’Université de Sherbrooke au Canada, fait état de résultats inédits jusqu’ici.
Les auteurs concluent à la nécessité de poursuivre les analyses d’impact des organismes génétiquement modifiés (OGM) sur les humains. Professeur-chercheur au département de gynécologie-obstétrique de la faculté de médecine de l’Université de Sherbrooke, Aziz Aris s’intéresse particulièrement à la toxicité d’origine environnementale sur les fœtus et les mères. Or, dit-il, l’impact des OGM a été abondamment testé sur les animaux, mais jamais sur les humains. C’est la première fois que la présence de résidus de pesticides d’OGM dans le sang des femmes est démontrée. Ces femmes et leur conjoint, étant citadins et n’ayant jamais travaillé au contact de pesticides, ces résultats résulteraient donc principalement de leur alimentation.
Les résultats de l’étude, financée par un fonds québécois de recherche en santé, seront publiés dans la revue scientifique américaine Reproductive Toxicology.
Environ 70% du maïs au Québec est génétiquement modifié (OGM) et inclut la toxine Bt.
Cette étude scientifique indépendante confirme les pires craintes de Greenpeace sur le manque de rigueur, d’indépendance et de transparence des études menées par les entreprises de biotechnologies.
Comme les autorités gouvernementales canadiennes refusent toujours d’appliquer le principe de précaution, les citoyens sont traités comme des cobayes par le gouvernement fédéral. Au Canada comme en Europe, les OGM sont toujours approuvés sans études sérieuses et indépendantes. Il est temps de sortir de l’obscurantisme !
Combien d’études scientifiques indépendantes seront nécessaires pour que les autorités gouvernementales exigent des entreprises de biotechnologies des preuves de l’innocuité des OGM ?
15:01 Publié dans Biologie, Environnement-Écologie, Mollusques, Nature et santé, Pollution | Tags : ogm, pesticides, femmes enceintes, foetus, glyphosate, glufosinate | Lien permanent | Commentaires (0) | Facebook | | |
05/03/2011
Cancers détectés par des chiens
Cancer de la prostate et cancers colorectaux détectés par des chiens
Étonnant : dans sa livraison de mars 2011, le mensuel Sciences et Avenir révèle que le flair du chien serait le meilleur outil à disposition des chercheurs pour progresser dans le dépistage de certains cancers. Dans le cas du cancer de la prostate, l'étude a été proposée à l'armée française par Olivier Cussenot, urologue à l'hôpital Tenon (Paris) et son équipe. Aspirant, un berger malinois de 2 ans et demi a été capable de détecter 30 malades sur 33 d'après leurs échantillons d'urine, soit un taux de réussite de 91 %. À comparer avec celui du test sanguin PSA appliqué à l'heure actuelle qui plafonne à 20 %... Deux ans d'apprentissage auront été nécessaires pour former Aspirant.
En ce qui concerne le cancer de la prostate, le test PSA (qui cible une protéine, la PSA, sécrétée par l'organe lors d'une inflammation) est tellement faillible qu'un dépistage systématique ne peut être envisagé. D'où l'intérêt d'un test plus spécifique. Les chercheurs ont engrangé plusieurs centaines de molécules candidates dans les urines de patients malades. « Tout l'intérêt de cette approche, explique Olivier Cussenot, consiste à accélérer l'identificalion. D'ici à deux ans, je pense qu'Aspirant nous aura permis de mettre la main sur plusieurs molécules nous conduisant à la mise au point d'un bien meilleur test. »
Ainsi, le chien se révèle un auxiliaire précieux en cancérologie. Une étude japonaise, parue Le British Médical Journal, publie une étude japonaise qui montre que le flair d'un labrador est plus efficace que la coloscopie pour la détection de cancers colorectaux dans les échantillons de selles de malades.
Source : Sciences et Avenir, n° 769 mars 2011 p. 39.
16:26 Publié dans Biologie, Médecine, Nature et santé, Poissons, Pollution | Tags : cancer de la prostate, cancer colorectal, détection, psa, détection par le chien | Lien permanent | Commentaires (3) | Facebook | | |
21/06/2010
La pomme de terre OGM "Amflora" interdite au Luxembourg
Le Luxembourg interdit la pomme de terre OGM "Amflora" !
par Greenpeace Luxembourg
Le 16 juin 2010, lors du colloque "La sécurité dans mon assiette" sur la sécurité de la chaîne alimentaire, le Ministre de la Santé luxembourgeois Mars Di Bartolomeo, en présence de Romain Schneider ministre de l’Agriculture, a interdit la culture de la pomme de terre OGM Amflora de BASF au Luxembourg.

L’interdiction de la pomme de terre OGM au Luxembourg a été annoncée quelques mois à peine après son autorisation par la Commission européenne et ceci devant des hauts représentants de l’autorité en charge de la sécurité alimentaire en Europe (EFSA).
C’est d’ailleurs au même endroit (Centre Culturel de rencontre Abbaye de Neumünster) que le Luxembourg avait déjà interdit le maïs transgénique MON810 en 2009.
Le message politique est courageux et sans appel possible : Le Luxembourg clame haut et fort qu’il n’est pas prêt à se laisser dicter un menu OGM par la Commission européenne. Après l’Autriche, c’est le deuxième état membre de l’Union européenne à prendre la décision de protéger les consommateurs et l’agriculture des conséquences négatives d’Amflora.
Greenpeace et les organisations partenaires de l’Initiative Luxembourg sans OGM (1) félicitent le Ministre de la Santé d’avoir pris ses responsabilités dans l’intérêt du consommateur et encouragent le Gouvernement à continuer de s’opposer résolument à la politique pro-OGM de la Commission européenne.
Une procédure d’évaluation qui doit être profondément réformée en Europe
Le 2 mars 2010, la Commission européenne avait autorisé la culture, l’utilisation dans les matières fourragères et la présence jusqu’à 0,9% dans l’alimentation humaine de la pomme de terre OGM Amflora développée par le géant allemand de la chimie BASF. Cette décision avait été prise malgré les évidences scientifiques relatives au risque de résistances aux antibiotiques.
Greenpeace et les organisations de l’initiative Luxembourg sans OGM tiennent à rappeler que procédure d’évaluation et d’autorisation des OGM dans l’Union européenne est défaillante et doit être réformée.
Les effets à long terme des OGM sur l’environnement et sur la santé, ainsi que leurs coûts socio-économiques pour toute la filière agricole ne sont actuellement pas pris en compte. « Nous demandons au Gouvernement luxembourgeois de continuer son engagement en faveur réforme de la une procédure d’évaluation et d’autorisation des OGM qui protège directement au niveau européen les consommateurs, l’environnement et l’agriculture des effets négatifs des OGM », déclare Maurice Losch de Greenpeace Luxembourg.
(1) : Les 29 organisations de l’Initative "Luxembourg sans OGM" :
Aide à l’Enfance de l’Inde, Association Solidarité Luxembourg Nicaragua, ASTM, Attac Luxembourg, Bauerenallianz, bioLABEL Lëtzebuerg, BIONA, Caritas Luxembourg, Demeter Bond Lëtzebuerg, Église Catholique de Luxembourg, Ëmweltberodung Lëtzebuerg asbl. (EBL), FCPT – SYPROLUX, FNCTTFEL – Landesverband, Fondation Hëllef fir d’Natur, Frères des Hommes, GREENPEACE Luxembourg, Haus vun der Natur, Initiativ Liewensufank, LCGB, Lëtzebuerger Natur- a Vulleschutzliga, Lëtzebuerger Landesverband fir Beienzuucht, Life asbl., Mouvement Écologique, OGBL, Slow Food Lëtzebuerg, SOS Faim Luxembourg, Stroossekanner Sao Paulo a.s.b.l., TransFair – Minka a.s.b.l., Union Luxembourgeoise des Consommateurs (ULC).
17:22 Publié dans Biologie, Environnement-Écologie, Nature et santé | Tags : greenpeace, ogm, santé, pomme de terre, luxembourg | Lien permanent | Commentaires (0) | Facebook | | |
08/12/2009
Gènes, pouvoirs et profits
Gènes, pouvoirs et profits
L'ouvrage "Gènes, pouvoirs et profits" de Christophe Bonneuil et Frédéric Thomas est maintenant disponible, en co-éditions Quae (INRA)/ FPH.
Ce livre retrace l'histoire des transformations conjointes de la génétique et de la société depuis cent cinquante ans. Trois grands régimes de production des savoirs et des innovations émergent. Au régime de la semence domestique succède après la seconde guerre mondiale le régime du progrès génétique planifié ; chercheurs et fonctionnaires, en lien avec la profession, produisent et régulent l'innovation variétale pour moderniser la « ferme France ». Ces régulations cèdent ensuite la place au régime contemporain de profit génétique mondialisé, creuset des OGM. Cet ouvrage s'adresse à ceux que l'avenir des semences, de l'alimentation et de la biodiversité préoccupent.
Pour plus d'information sur cet ouvrage ou pour passer une commande :
http://www.quae.com/livre/?gcoi=27380100316660&source=alerts&utm_medium=emailalerts&utm_campaign=alerts
Broché, 624 p., 55 € + 5 € de frais de port.
07:30 Publié dans Actualité des Sciences, Biologie | Tags : science, génétique, agriculture, semences, ogm | Lien permanent | Commentaires (0) | Facebook | | |
18/11/2009
L'Escargot (1) : Morphologie externe
L'Escargot, cet animal hermaphrodite, est un excellent modèle pour l'étude de la sexualité et le déterminisme du sexe. En effet, les cellules souches de sa gonade évoluent soit en ovules, soit en spermatozoïdes. Voici toute une série d'articles sur cet animal fabuleux. Ce premier chapitre traite l'étude de la morphologie de l'animal.
10:22 Publié dans Biologie, Invertébrés, Mollusques, Sexualité-embryologie | Tags : sexualité, hermaphrodisme, escargot, mollusques, morphologie | Lien permanent | Commentaires (0) | Facebook | | |
17/11/2009
L'Escargot (2) : La coquille
Dans ce deuxième chapitre consacré à l'Escargot, intéressons-nous à la structure de la coquille, à sa régénération en cas de fracture et ses applications en cosmétologie
10:42 Publié dans Biologie, Invertébrés, Mollusques, Sexualité-embryologie | Tags : escargot, coquille, mollusques, gastéropodes | Lien permanent | Commentaires (0) | Facebook | | |