



11/10/2009
Alzheimer : ça se précise
Alzheimer : ça se précise
par Jean-Jacques Perrier (journaliste à "Pour la Science")
Deux études génétiques de grande envergure précisent le mécanisme hypothétique de la maladie d'Alzheimer.
Pour mieux comprendre les causes de la maladie d'Alzheimer, les chercheurs réalisent des études génétiques dites d'association. Ils comparent des milliers de mutations ponctuelles d'ADN variant entre individus, des marqueurs nommés SNP, chez un grand nombre de malades et de personnes en bonne santé. Dans la forme sporadique de la maladie d'Alzheimer, qui représente 97 pour cent des cas, le seul facteur génétique de prédisposition confirmé dans toutes les études a été découvert en 1993 : il s'agit d'une forme du gène codant l'apoliprotéine E (ApoE), l'allèle epsilon 4. Depuis, rien n'a vraiment émergé de concluant… jusque récemment.
Grâce à des études dites pangénomiques, qui analysent les 23 paires de chromosomes dans leur ensemble, deux consortiums internationaux ont analysé et comparé près de 600 000 marqueurs chez plus de 20 000 personnes. Le travail coordonné par Philippe Amouyel, à l'Institut Pasteur de Lille (Inserm U744), révèle l'implication de deux gènes, celui de la clusterine et celui codant une protéine contribuant au système immunitaire, CR1 (récepteur 1 du composant 3b/4b du complément). Et l'étude coordonnée par Julie Williams, à l'Université de Cardiff, montre une association avec le gène de la clusterine, là encore, et avec le gène codant la protéine PICALM (phosphatidylinositol-binding clathrin assembly protein).
Des résultats précédents, et d'autres en cours de publication, suggèrent que les deux premières protéines (la clusterine et CR1) interviennent, avec l'ApoE, dans l'élimination du peptide bêta-amyloïde (PICALM est quant à elle impliquée dans le fonctionnement des synapses, les connexions entre neurones). Or, selon l'explication la plus commune de la maladie, l'accumulation du peptide bêta-amyloïde provoque l'une des lésions cérébrales caractéristiques : les plaques séniles (voir Pour oublier la maladie d'Alzheimer, Pour la Science, août 2006). La forme tardive d'Alzheimer serait ainsi associée à une élimination insuffisante de cette molécule du cerveau, tandis que les formes héréditaires, plus précoces, seraient liées à sa surproduction.
22:40 Publié dans Médecine, Nature et santé | Tags : santé, alzheimer | Lien permanent | Commentaires (0) |  Facebook | | |
Facebook | | |
10/10/2009
La Roche de Solutré et son musée préhistorique
 La Roche de Solutré et son musée préhistorique
La Roche de Solutré et son musée préhistorique
par André Guyard
La Roche de Solutré est un site rendu célèbre par l'ascension annuelle qu'entreprenaient chaque lundi de Pentecôte, François Mitterrand et ses fidèles.
Le site de Solutré est sans conteste un des hauts lieux de la Bourgogne méridionale. Il est remarquable par sa composition paysagère insolite en Bourgogne. Cet éperon rocheux dominant le village de Solutré-Pouilly et le vignoble environnant a fait l'objet depuis plus d'un siècle de mesures de protection. Ensemble patrimonial prestigieux, il est exceptionnel à plus d'un titre.
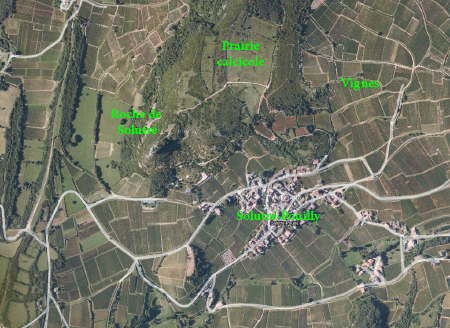
Géologie et paysage
Le Grand Site de Solutré-Pouilly-Vergisson, qui constitue aujourd'hui une entité paysagère et humaine à part entière, doit son caractère spectaculaire à son histoire géologique.
Ce paysage original est remarquable, en particulier, par le rocher dressé au-dessus du vignoble du Maconnais. L'explication est à la fois géologique et tectonique : nous sommes, en effet, sur la bordure sud-orientale du Bassin parisien bouscule et fracture lors du plissement alpin. Les couches calcaires déposées dans les mers chaudes de l'ère secondaire ont été soulevées et fracturées ; l'érosion a, par la suite, mis à profit cette vulnérabilité pour dégager les matériaux les plus tendres et laisser en évidence des abrupts de calcaires coralliens. Ce phénomène est particulièrement mis en évidence ici où la dissymétrie du relief est évidente : la masse calcaire formée de plusieurs bancs superposés présente un front abrupt tourné vers le sud-ouest alors que son revers s'incline doucement vers le nord-est.

Ce lambeau de plateau, situé à environ 450 m d'altitude porte les traces d'une ancienne exploitation agricole ou pastorale sous la forme d'alignements d'épierrement soulignés par des haies : ces terres plutôt ingrates sont aujourd'hui abandonnées à de maigres boisements ou aux pelouses calcicoles ; la grande parcelle rectangulaire sur la droite du rocher offre une surface appréciable de ce milieu particulier.
Outre le vignoble qui entoure la roche de toute part, on distingue sur ce modeste territoire, d'autres unités caractéristiques : les roches, les pelouses sèches, la forêt et le bocage. Le contraste entre les falaises abruptes et cette diversité dans la lecture des panoramas est tout à fait singulier en Bourgogne méridionale.

Un site préhistorique
Depuis le XIXe siècle, Solutré occupe une place éminente dans la Préhistoire française. En 1866, un site archéologique majeur a été découvert au pied de la Roche de Solutré. Ont été mis au jour d'impressionnants gisements d'ossements animaux et des outils du Paléolithique supérieur. Les nombreuses fouilles qui s'ensuivront, facteur essentiel des premières mesures de protection, et les découvertes qui en seront tirées feront de Solutré un site archéologique de premier plan en Europe. Solutré, par la richesse des découvertes, devint même le site éponyme d'une culture du Paléolithique supérieur : le Solutréen (de 20 000 à 16 000 ans avant J.C.) caractérisée par ses "feuilles de laurier", chef d'œuvre de la taille du silex.
Parmi les trouvailles, de nombreux objets ont été regroupés dans un remarquable musée souterrain situé au pied de l'abrupt de l'éperon rocheux.
Ce qui fait surtout l'originalité de ce musée, c'est la reconstitution de l'élaboration de ces silex taillés en feuille de laurier. Un travail méticuleux qui demandait une haute technicité à l'artisan paléolithique.













Une biodiversité remarquable à préserver

Ce site est par ailleurs remarquable par le biotope que constituent les pelouses calcaires présentes au sommet des roches. Grâce à des conditions particulières de sols et d'exposition, les pelouses abritent des plantes "méditerranéo-montagnardes" éloignées de leur station d'origine, telles l'Inule des montagnes, le Coronille arbrisseau, le Micrope droit... On note également une grande richesse en orchidées dont certaines très rares.
La faune comporte notamment des espèces d'oiseaux inféodées aux pelouses sèches (Bruant ortolan et Hibou Petit Duc) et une population de rapaces spécialistes des éboulis et des falaises.
En reconnaissance de l'intérêt européen de son patrimoine naturel, ce site a été intégré au réseau européen Natura 2000.
Un site menacé
Bien que bénéficiant d'un classement au titre des monuments naturels (loi de 1930), d'un arrêté de protection de biotope depuis 1991 et officiellement devenu Grand Site en décembre 2000, ce site est pourtant en danger : en l'absence de gestion coordonnée et durable, la banalisation du paysage et ['importance de sa fréquentation mettent en péril son caractère remarquable. Par exemple sur les pelouses, l'abandon des pratiques de pâturage laisse la voie à une fermeture du milieu, sans intervention l'embuissonnement menace et la qualité des paysages risque par processus spontané de végétalisation de se voir banalisé.
Des actions en faveur de la préservation de ces milieux naturels
Dans le cadre d'une convention de gestion avec le Syndicat mixte de Valorisation du Grand Site de Solutré-Pouilly-Vergisson, le Conservatoire des Sites Naturels Bourguignons intervient depuis de nombreuses années sur ce site : en 2002 un troupeau de Konik Polski est installé pour la mise en œuvre d'un pâturage écologique. Depuis l'équipe du Conservatoire intervient régulièrement pour assurer un suivi du cheptel, pour la pose de clôture, pour effectuer des suivis scientifiques. En 2007 le plan de pâturage a été réactualisé, les pelouses du mont de Pouilly ayant été intégrées à la gestion tournante des pelouses, le calendrier de pâturage a été redéfini et un suivi scientifique de l'impact du pâturage a également été conduit. Depuis 2009, des panonceaux d'information sur les koniks Polskis sensibilisent les visiteurs.

Les roches de Solutré et Vergisson sont devenues au fil des ans le symbole emblématique de la région Bourgogne et plus particulièrement de la Bourgogne du sud à la fois par la beauté de leurs paysages et par la richesse patrimoniale et culturelle qui y sont rattachées. Le Syndicat mixte du Grand Site s'attache à développer, dans le cadre de l'Opération Grand Site, des actions pour sa gestion durable tout en continuant à accueillir des visiteurs de plus en plus nombreux.
Sources :
- Solutré-Pouilly-Vergisson, Bourgogne Nature n° 8, 2008.
- Documents photographiques : André Guyard
07:42 Publié dans Préhistoire | Tags : solutré, solutréen, préhistoire, paléolithique | Lien permanent | Commentaires (1) | Facebook | | |
09/10/2009
Le Bihoreau gris
 Le Bihoreau gris
Le Bihoreau gris
(Nycticorax nycticorax)
par André Guyard
Le Bihoreau gris est un oiseau trapu avec une tête large, un cou épais et court, et des pattes courtes.
En période de reproduction, l'adulte montre deux longues plumes blanches disposées sur la nuque.

Le Héron bihoreau est habituellement silencieux. Il émet quelques cris en volant ou depuis un perchoir.
Le Héron bihoreau vit près des lacs, des marécages et des rivières bordés de végétation dense. Il niche et dort dans les arbres.
C'est un oiseau nocturne, se nourrissant du crépuscule à l'aube. Le Héron bihoreau se nourrit principalement de poissons, mais aussi de vers de terre, et d'insectes aquatiques et terrestres. Comme les autres hérons, il chasse dans les eaux peu profondes, utilisant son long bec pour capturer les proies. À l'affût, il se tient debout immobile, attendant le passage d'une proie. Il secoue vigoureusement sa proie pour l'étourdir ou la tuer et il l'avale la tête la première.

Le Bihoreau gris
Le vol est direct marqué par des battements assez rapides et peu amples. Au vol, le bec est incliné vers le bas. Seuls les doigts dépassent la queue.
Le Héron bihoreau niche en colonies souvent avec des espèces voisines dans les fourrés, sur les arbres et localement dans les roseaux. Le mâle bâtit la base du nid le plus souvent dans les branches d'un buisson ou au faîte d'un arbre où il attirera la femelle par des mouvements et des cris particuliers. C'est la femelle qui se chargera de terminer le nid.
La femelle dépose 3 à 5 œufs bleu clair, à intervalles de deux jours. L'incubation dure environ 24 à 26 jours, assurée par les deux parents.
Les poussins sont couvés par les deux adultes et nourris par régurgitation. Ils quittent le nid au bout de trois semaines.
Vidéo : André Guyard
Pour en savoir plus : http://www.oiseaux.net/oiseaux/bihoreau.gris.html
13:35 Publié dans Ornithologie | Lien permanent | Commentaires (0) | Facebook | | |
07/10/2009
Nouvelles pistes de dinosaures découvertes dans l'Ain
 Nouvelles pistes de dinosaures
Nouvelles pistes de dinosaures
découvertes dans l'Ain
par André Guyard
(Dernière mise à jour : 02/02/2018)
(Document BRGM)

Après Coisia, Loulle et Courtedoux, de nouvelles pistes de dinosaures ont été découvertes dans le massif jurassien, sur le plateau de Plagne dans l'Ain, à près de 800 m d'altitude. Ce sont les plus grandes empreintes jamais mises à jour : certaines atteignent 1,50 m de diamètre. Cette trouvaille a été réalisée en avril 2009 par deux naturalistes amateurs : Marie-Hélène Marcaud et Patrice Landry qui font partie de la Société des Naturalistes d'Oyonnax. Rappelons que les géologues de la SDNO sont déjà à l'origine de la découverte du site de Coisia. Alertés, les paléontologues, Pierre Hantzpergue du laboratoire Paléoenvironnements et paléobiosphères de l'Université de Lyon-1 et Jean-Michel Mazin du CNRS ont expertisé le site.
À Plagne, ce sauropode de 40 t a laissé une suite de pas de 155 m. Après deux ans d'analyses, l'animal du jurassique vient de livrer son identité.

La piste d'empreintes de sauropode de Plagne
© P. Dumas
Depuis 150 millions d'années, ces empreintes ont été conservées dans une strate calcaire, représentant un sédiment lagunaire pétrifié. Sur le cliché ci-dessous apparaît magnifiquement dessinée, l'empreinte d'un pied dans le sédiment, bordée d'un bourrelet de boue pétrifiée. Le témoin de 50 cm indique que la taille de l'empreinte mesure approximativement 150 cm.

Un beau coup de patte !
(Cliché Pierre Hantzpergue)
Comment se forment les empreintes
Pour que des empreintes laissées au bord de l'eau se conservent des millions d'années, les conditions de sédimentation doivent être idéales. « Le sol doit être compose de sédiments très fins, qu'il s'agisse de vase argileuse ou de calcaire », explique Pierre Hantzpergue (CNRS-université de Lyon-1) qui a étudié différents sites du Jura comme Plagne ou encore Loulle où 1500 empreintes ont aussi été retrouvées. «Il faut un compromis entre la finesse, qui permet le détail, et une consistance pas trop boueuse pour que l'animal ne dérape pas », précise Nicolas Olivier (université Clermont-Auvergne-CNRS). Cette vase à la plasticité d'une pâte à modeler, doit ensuite sécher rapidement, ce qui suppose un climat chaud. Aussi le scénario est-il souvent le suivant : à marée basse, des dinosaures se promenant sur l'étendue boueuse du rivage laissent leurs empreintes... que la mer vient recouvrir, déposant une microfiche de sédiments.
La sédimentation marine a ensuite accumulé des centaines de mètres de sédiments protégeant ainsi ces strates pendant des millions d'années. Le Jura ayant émergé, l'érosion s'est chargée du déblaiement des couches supérieures. C'est ainsi que les traces de ces sauropodes ont été mises à jour par le passage d'engins forestiers qui ont décapé la mince couche de terre végétale qui dissimulait la dalle.
Comme à Loulle ou à Coisia (voir articles correspondants à ces deux localités jurassiennes) il s'agit de sauropodes, des dinosaures herbivores au long cou. D'après Pierre Hantzpergue, on ne trouve pas de traces de pas de cette taille nulle part ailleurs. Elles correspondent à des animaux de 25 m de long pesant 30 à 40 tonnes.
Certes, il ne s'agit pas d'Amphicoelias fragillimus, le plus grand de tous les dinosaures trouvés jusqu'à présent qui atteint une longueur de 40 à 50 m pour un poids estimé à 120 tonnes et qui vivait en Amérique du Nord également au jurassique (du Kimméridgien au Tithonien entre 155 et 154 millions d'années), mais les empreintes laissées par ces mastodontes sont les plus grandes jamais observées en France.
Il y a 150 millions d'années, le Jura ressemblait davantage aux Bahamas qu'à la région montagneuse d'aujourd'hui. Grâce à ce lointain passé, Plagne, village peu connu du grand public situé au sud de la chaîne du Jura, près d'Oyonnax (Ain), abrite des témoignages parmi les plus fascinants de l'univers disparu du jurassique. Et les vestiges préservés dans le sol, miraculeusement exhumés, pourraient relancer la « dinomania » des années 1990.
Découverte la plus spectaculaire :
Cette suite de pas de 155 mètres, record du monde de la plus longue piste d'empreintes de sauropodes les plus imposants des dinosaures. Après trois campagnes de fouilles entre 2010 et 2012, le paléontologue Jean-Michel Mazin et le spécialiste de paléoenvironnement Pierre Hantzpergue, du Laboratoire de géologie de Lyon, et Nicolas Olivier du Laboratoire Magmas et Volcans de Clermont-Ferrand, ont publié leurs résultats en novembre 2017 dans Geobios. Ils dressent le portrait d'un animal pesant au moins 40 tonnes pour 35 à 40 mètres de long se promenant au milieu des fougères, des mousses et des conifères. Dans cette immense lagune — une plateforme carbonatée qui se soulèvera bien plus tard lors de la formation des Alpes —, des îles émergeaient d'une mer chaude et peu profonde. C'est là que s'ébattait le reptile géant.

"Une piste de 155 mètres a été dégagée" précise Pierre Hantzpergue qui a commencé les fouilles. Une perspective prometteuse qui pourrait faire de Plagne le plus grand site de pistes de dinosaures du monde. Les résultats des fouilles qui dureront au moins trois ans seront comparés avec ceux issus des sites de Coisia, Loulle et Courtedoux, ce dernier site situé dans le Jura suisse et permettront de reconstituer l'histoire de la région au jurassique, car ces pistes sont inscrites dans des terrains d'âge différant de plusieurs millions d'années, entre -155 et -150 millions d'années.
L'identité du monstre enfin révélée
Après deux ans d'analyses, l'animal vient de trouver une identité : Brontopodus plagnensis, une ichno-espèce (décrite uniquement par son empreinte) appartenant au groupe des titanosaures, les plus lourds ayant jamais existé, et ne possédant pas de correspondant connu chez les fossiles. Ses empreintes prennent la forme de dépressions rondes ou ovales, entourées d'un bourrelet de sédiments calcaires expulsés par le poids de l'animal en mouvement. « Plus une piste est longue et plus elle raconte une histoire, se réjouit Jean-Michel Mazin. Ainsi, 155 mètres c'est 3 minutes et demie dans la vie de l'animal. Nous avons dégagé 115 pas environ. On le voit ainsi changer de direction, accélérer en passant de 3,5 à 4,5 km/h. Au début de son cheminement, ses mains — comme on appelle ses petites pattes de devant — et ses pieds — ses pattes arrière — sont bien séparées. Puis il allonge le pas et le pied écrase l'empreinte de la main ! » Avec des enjambées pouvant atteindre 2,80 m... Non content de battre le record de longueur de piste, auparavant détenu par le site de Galinha au Portugal (147 mètres), Brontopodus plagnensis grimpe aussi vers les sommets de l'échelle des pointures : selon la nature du sol et les bourrelets formés, ses traces mesurent de 96 à 112 cm. Ces marques de pied découvertes à Plagne (Ain) sont celles d'un « Brontopodus plagnensis » pesant au moins 40 tonnes. Elles mesurent de 96 à 103 cm. Insuffisant cependant pour battre le record détenu par un sauropode australien doté d'un pied de 170 cm.

Sauropode de Plagne marchant sur ses empreintes ;
© dessin : A. Bénéteau ; photographie : Dinojura
(Cliquer sur le cliché pour l'agrandir)
BIOMÉCANiQUE
4 km/h, la vitesse moyenne des dinosaures
Retrouver la vitesse de déplacement d'un animal à partir de ses enjambées est possible grâce à une formule mathématique mise au point en 1975 par Robert McNeill Alexander, zoologiste britannique spécialisé dans la biomécanique et la locomotion terrestre des mammifères :
v= 0,25g-0,5S1,67H-1,17,
g étant l'accélération de la pesanteur au sol exprimée en mètres par seconde au carré (9,81 m/s2), S la longueur de l'enjambée (distance entre deux empreintes de pied du même côté) et h la hauteur à la hanche de l'animal.
« Cette dernière n'est pas mesurable sur une piste, mais à partir de squelettes fossiles, explique le paléontologue Jean-Michel Mazin. Nous avons établi une relation entre longueur du pied et hauteur à la hanche des différents types de dinosaures. La hauteur varie de 4,5 à 5,9 fois la longueur du pied.» Un calcul a montré que les grands sauropodes ne se déplaçaient pas à plus de 4 km/h. Le chercheur et ses collègues expérimentent régulièrement cette formule avec les enfants venus visiter Crayssac. Ils les font marcher sur un sol qui imprime leurs empreintes, prennent leur hauteur de hanche et les résultats correspondent à la formule !

Le site ne recèle "pas d'ossements et on n'en trouvera pas parce que ces empreintes de pas se conservent dans des environnements qui ne permettent pas la conservation des squelettes. Pour qu'il y ait préservation d'un squelette, il faut que ce soit dans l'eau rapidement recouverte par des sédiments", précise M. Hantzpergue.
Les fouilles à venir dégageront une piste le plus loin possible, ainsi qu'un carré d'un demi-hectare à un hectare, pour déterminer la densité des traces.
Ces travaux "pourraient révéler que le site de Plagne est l'un des plus vastes connus au monde", estime le CNRS. Ainsi le département de l'Ain détient donc aujourd'hui un record du monde, celui de la piste d'empreintes la plus longue, un chemin de dinosaures plus long que la plus grande piste connue aujour'hui qui fait 147 mètres au Portugal, près de Fatima.
Sources :
Merci au professeur Pierre Hantzpergue qui nous a fourni les clichés qui illustrent cet article.
Science et Vie, n° 1106, novembre 2009, p. 17.
Voir également le site de la SDNO dédié à la découverte de Plagne.
Sciences et Avenir, n° 852, février 2018, pp. 56-58.
Futura Planète : Empreintes de dinosaures : la plus longue piste de sauropode est en France !
17:44 Publié dans Actualité des Sciences, Paléontologie - archéologie | Tags : dinosaures, sauropodes, fossiles, pistes, jurassique, jura, brontopodus plagnensis, plagne | Lien permanent | Commentaires (0) | Facebook | | |
Voie lactée : son recensement est enfin terminé
 Voie lactée : son recensement est enfin terminé
Voie lactée : son recensement est enfin terminé
par André Guyard

Le Terre et la Voie lactée
LA VOIE LACTÉE
La voie lactée est une galaxie spirale géante, comme il en existe des dizaines de milliards dans l'Univers visible, il s'agit d'un vaste disque en rotation, renflé en son centre, composé d'étoiles et de gaz interstellaire. Autour de ce disque, un immense halo, peuplé de très vieilles étoiles, constitue le vestige de la formation de cette vaste agglomération stellaire, voici 13 milliards d'années. La Voie lactée mesure environ 100 000 années-lumière de diamètre. Le système solaire se trouve dans le disque de la Voie lactée, à 28 000 années-lumière du noyau central.


La Voie lactée est un élément important du "puzzle" cosmologique : comprendre comment elle est apparue, quelques centaines de millions d'années après le big bang et comment elle a évolué, c'est appréhender l'évolution de l'Univers dans son ensemble. Afin d'étudier globalement la Voie lactée, une équipe française a créé un outil unique, fusionnant observations télescopiques et calculs théoriques, le "modèle standard".
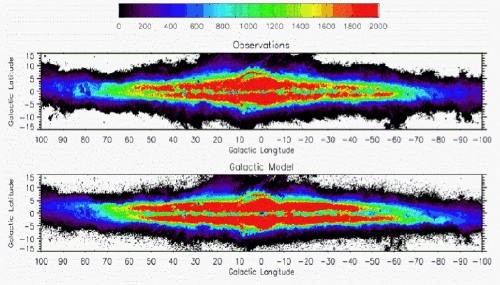
C'est le plus formidable recensement céleste jamais réalisé par des astronomes. Entrepris par une équipe de l'observatoire de Besançon voici un quart de siècle, il livre aujourd'hui un résultat vertigineux : la première estimation précise de la population stellaire de notre galaxie, la Voie lactée...
À l'origine de ce projet unique, entamé en 1985, les astronomes français Annie Robin et Michel Crézé, rejoints au tournant du millénaire par Céline Reylé et Mathias Schultheis. Le résultat ? 140 milliards d'étoiles, plus ou moins dix milliards. Une première : jusqu'ici, les populations stellaires de la Voie lactée n'étaient que grossièrement estimées. Désormais, l'équipe d'Annie Robin peut discriminer avec précision les différentes populations et générations d'étoiles naines, géantes, jeunes, vieilles, etc.
UNE TÂCHE HERCULÉENNE
Si ce dénombrement a attendu aussi longtemps pour être délivré, c'est que le recensement de la Voie lactée était une tâche herculéenne. D'abord, parce que le système solaire où nous habitons se trouve plongé au sein même de ce vaste disque stellaire, renflé en son centre. Or, il est malaisé d'étudier un système depuis l'intérieur, surtout qu'une grande partie de la Galaxie nous est masquée par l'accumulation de gaz et de poussières interstellaires présents dans son disque, qui agissent comme un brouillard dense et impénétrable !
Ensuite, les dimensions de notre galaxie sont telles que même avec le télescope spatial Hubble ou les télescopes géants terrestres, il est impossible de détecter tous les astres qui la composent... Ainsi, notre image de la Voie lactée restait-elle très parcellaire. Pour preuve, le plus grand catalogue astronomique actuel, le catalogue USNO-B1.0, dressé en un demi-siècle d'observation continue par les grands observatoires du monde ne contient "que".... un milliard d'astres, soit moins de 1 % du chiffre annoncé par le groupe de Besançon !
Mais l'équipe d'Annie Robin, directeur de recherche au CNRS, a trouvé une solution aussi efficace qu'élégante pour contourner ces obstacles: mixer observations et calcul statistique. "Nous utilisons les observations astronomiques, menées sur des champs limités du ciel, dont nous projetons ensuite les propriétés sur l'ensemble de la Galaxie ", explique-t-elle. Ou comment les parties révèlent in fine le tout ! En effet, la Voie lactée, que l'on peut assimiler à un disque en rotation, est un astre symétrique, homogène, dont la forme et la dynamique sont gouvernées par son seul champ de gravitation.


Dès lors, les "sondages profonds", ces images électroniques prises avec un très long temps de pose dans des régions soigneusement choisies de la Voie lactée, permettent, lorsqu'ils sont intégrés au calcul théorique, de dessiner le profil exact de notre gigantesque agglomération stellaire. Annie Robin poursuit: "À l'observatoire de Besançon, afin de mieux comprendre l'origine, l'évolution et la structure de la Voie lactée, nous avons créé l'équipe Galaxie. Notre groupe développe un modèle numérique de Voie lactée, à partir des observations réelles et des modèles de populations stellaires." Un travail de bénédictin, en constant renouvellement depuis vingt-cinq ans, qui a permis en 2003 la publication par Annie Robin, Cécile Reylé et leurs collaborateurs d'un "modèle standard" de notre galaxie, dit aussi "modèle de Besançon", actuellement unique au monde et auquel se réfèrent désormais tous les chercheurs qui étudient la Voie lactée. Concrètement, l'équipe a mis en ligne son modèle sous forme de programme interactif. Les astronomes professionnels peuvent étudier la Voie lactée en lançant ce programme après avoir préalablement choisi les paramètres stellaires qui les intéressent.
Le portrait brossé de la Voie lactée par le modèle de Besançon a apporté son lot de surprises, à commencer par le nombre d'astres recensés : "En fait, nous nous attendions à plus d'étoiles : la plupart des astronomes estimaient le nombre total d'étoiles entre 200 et 400 milliards... alors qu'il n'y en a que 140 milliards", souligne Céline Reylé. Ensuite, ce que révèle le modèle, c'est un extraordinaire déséquilibre entre les populations d'étoiles. Les petites étoiles de faible masse sont très largement majoritaires. Près de 60 % d'entre elles sont des naines rouges, cent à dix mille fois moins lumineuses que le Soleil. Par contraste, la population d'étoiles supergéantes, plus de vingt fois plus massives que le Soleil, est étonnamment restreinte, puisqu'elles représentent moins de 0,00001 % du total, soit quelques milliers d'astres !
Et le Soleil ? Celui-ci est souvent considéré, dans la littérature scientifique, comme une "étoile moyenne". En fait, il n'en est rien : le pourcentage des étoiles de masses comprises entre un demi et deux soleils est inférieur à seulement 15 % ! Quant aux étoiles rigoureusement semblables au Soleil, c'est-à-dire présentant la même masse, la même taille et le même éclat, elles ne sont "que" 2,4 milliards, ce qui ne représente que 1,7 % de la population galactique.
Un résultat vertigineux : 140 milliards d'étoiles, plus ou moins 10 milliards !
Tout compte fait, la Voie lactée n'accueille que cent milliards de véritables étoiles, à savoir des astres qui tirent leur énergie d'une réaction thermonucléaire. Les quarante milliards d'astres restants sont des "étoiles ratées" ou des vestiges d'étoiles mortes, pour la plupart invisibles car brillant d'un éclat extrêmement faible...
UN MODÈLE NUMÉRIQUE...
En tête arrivent les naines brunes : une population énorme (environ 30 milliards, soit 21 %) mais connue et étudiée depuis une quinzaine d'années seulement. Il s'agit d'étoiles trop peu massives pour que s'enclenchent en leur cœur des réactions nucléaires. Seule la pression gazeuse, qui les chauffe, les fait briller.
Puis viennent les naines blanches un peu plus de dix milliards d'individus, soit 7 % qui sont en fait les cœurs mis à nu d'anciennes supergéantes disparues après leur explosion.
Restent les étoiles à neutrons et les trous noirs, impression- nants vestiges des plus massives des supergéantes : les étoiles à neutrons ne sont que des blocs hyperdenses de particules, dont la taille ne dépasse pas... 20 km ! Il en existe un milliard environ dans la Galaxie, soit 0,7 %. Quant aux trous noirs, dont la masse, proche de celle du Soleil, est condensée dans le volume... d'une particule élémentaire, ils sont eux aussi environ un milliard |0,7 %) dans la Voie lactée.
À tous ces paramètres, il faut ajouter une inconnue... de taille, la fameuse matière noire, une matière invisible, transparente, indétectable et de nature mystérieuse, qui baignerait en théorie la Galaxie comme, d'ailleurs, toutes les galaxies de l'Univers. "La masse totale de la Voie lactée incluant la matière noire, atteint mille milliards de fois la masse du Soleil. En revanche, lorsque l'on prend en compte seulement les corps visibles — étoiles, nuages interstellaires, etc. —, la masse de la Voie lactée atteint seulement 50 milliards de masses solaires", explique Céline Reylé. "On a longtemps cherché à expliquer cette énorme masse de matière invisible par la présence dans la Galaxie d'astres indétectables, comme les naines brunes. Mais les observations ont permis d'éliminer cette hypothèse. Aujourd'hui, les physiciens cherchent l'explication à cette énigme du côté des particules exotiques, qui n'interagissent pratiquement pas avec la matière normale."
... ADOPTÉ PAR LES SCIENTIFIQUES
Le "modèle standard" de l'observatoire de Besançon est si précis et efficace dans sa capacité à corréler observation et théorie qu'il a été adopté par les astronomes européens engagés dans le projet de futur satellite astronomique Gaia. Il s'agit d'un télescope spatial de l'Agence spatiale européenne qui doit s'envoler de Kourou fin 2012.


© ESA, Gaia, DPAC La première carte de la voie lactée vue par Gaia
Sources :
— Article de Isabelle Brunnarius sur France 3 Franche-Comté
09:05 Publié dans Astronomie | Tags : voie lactée, dénombrement étoiles, gaia, observatoire de besançon | Lien permanent | Commentaires (0) | Facebook | | |
La Voie lactée étend ses bras
 La Voie lactée étend ses bras
La Voie lactée étend ses bras
Des observations récentes révèlent que l'un des bras spiraux de notre galaxie est plus étendu qu'on ne le pensait (voir l'article Voie lactée : son recensement est terminé).
De l'avis de la plupart des astronomes, la Voie lactée est une galaxie spirale barrée comportant deux bras principaux, celui de Persée et celui de l'Écu-Croix (voir l'illustration ci-dessous). Les autres bras seraient des structures mineures, faites surtout de gaz. Thomas Dame et Patrick Thaddeus, astrophysiciens à Harvard, aux États-Unis ont découvert ce qui est probablement un prolongement du bras de l'Écu-Croix, bras qui entoure désormais presque toute notre galaxie, depuis sa barre centrale d'étoiles jusqu'à son bord. Le bras de l'Écu-Croix apparaît ainsi comme le symétrique du bras de Persée, ce qui confère à la Voie lactée une structure plus simple et plus symétrique qu'on ne le croyait.
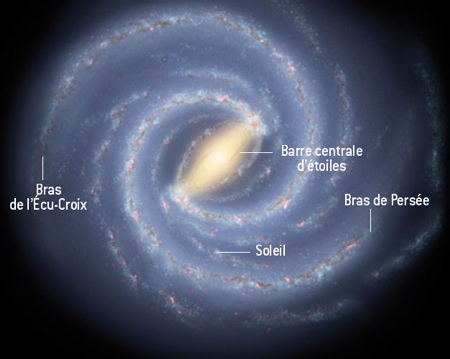
Vue d’artiste de la Voie lactée, qui intègre la découverte
du prolongement du bras de l’Écu-Croix
Le nouveau bras ou morceau de bras serait long d'environ 60 000 années-lumière, alors que le diamètre de la Voie lactée en fait 100 000. Pourquoi n'avait-il pas été remarqué auparavant ? Probablement parce qu'il est courbé : il se trouve en effet légèrement au-dessus du disque voilé de notre galaxie. Il aurait ainsi échappé à l'attention des astronomes, qui l'avaient déjà observé sans l'identifier.
Les deux astrophysiciens l'ont découvert en tentant de suivre les bords du bras de l'Écu-Croix. À leur grande surprise, ils ont détecté une structure surgissant à un angle de trois degrés au-dessus du plan galactique, dans le prolongement linéaire de l'Écu-Croix. Afin de s'assurer de leur découverte, ils ont examiné les données préexistantes d'une cartographie de l'hydrogène neutre dans notre galaxie et ses alentours. Cela leur a permis de tracer l'aspect du bras tel qu'il est observé à la longueur d'onde de 21 centimètres.
Puis ils ont eux-mêmes réalisé des observations dans le domaine millimétrique pour détecter la présence de monoxyde de carbone (CO) à une dizaine d'endroits le long du nouveau bras. Cette présence est révélatrice de nuages denses d'hydrogène moléculaire où se forment les étoiles, nuages qui constituent les meilleurs traceurs des bras spiraux. Ils démontrent ainsi que le nouveau bras n'est pas une illusion.
© ESA, Gaia, DPAC La première carte de la voie lactée vue par Gaia
Pour en savoir plus :
— Article de Isabelle Brunnarius sur France 3 Franche-Comté
— Pour la Science, juillet 2011.
T. M. Dame and P. Thaddeus (2011). - A molecular spiral arm in the far outer galaxy Astrophysics J. Letters, 734
http://arxiv.org/abs/1105.2523
08:32 Publié dans Astronomie | Tags : voie lactée, bras spiraux | Lien permanent | Commentaires (0) | Facebook | | |
Champignons coprophiles
 Champignons coprophiles
Champignons coprophiles
Quand l’esthétisme se cache dans la crotte
par André Guyard
Les champignons coprophiles sont des champignons qui croissent sur les excréments animaux.
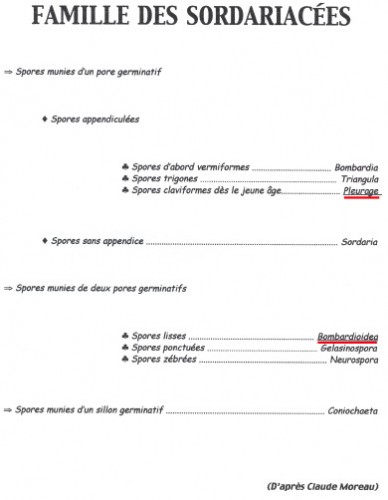
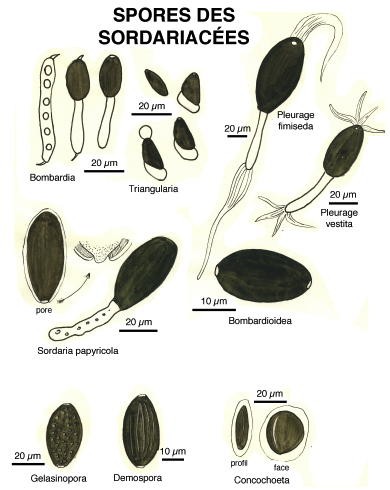
Pour la plupart, ce sont des Ascomycètes, c’est-à-dire des champignons dont les spores sont formées dans des espèces de sacs ou périthèces remplis d’un bouquet d’utricules appelés asques. Les périthèces émergent hors du support et, phénomène étonnant, à maturation, les ascospores vont être disséminées par une projection hors des asques, comme, par exemple, dans la famille des Ascobolacées.
Au cours de l’année de formation professionnelle à l’école normale, chaque élève-maître devait fournir une monographie. André Bride, professeur de mathématiques et éminent mycologue m’avait trouvé un sujet : les champignons coprophiles.
C’est ainsi que je passai mon automne 1958 à récolter crottes de lapins de garenne et de lièvres, à les observer sous loupe binoculaire, à les dessiner à une chambre claire et à les déterminer avec l’aide de mon mentor. Dessins au crayon, encres de Chine, aquarelles et gouaches : des documents cinquantenaires et… inédits.
Ci-dessous, quelques extraits de cette étude.




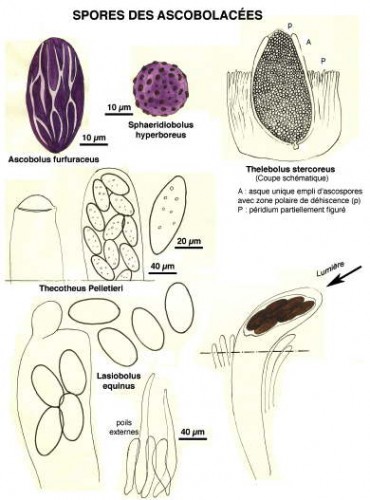
Dessins de André Guyard (1959)
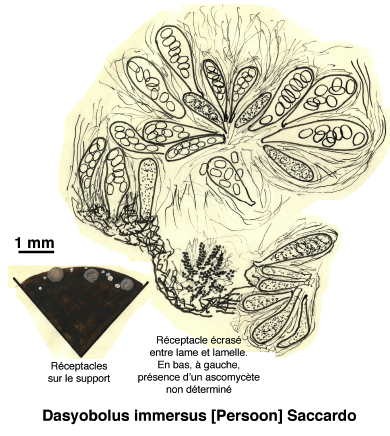
Dessins de André Guyard (1959)
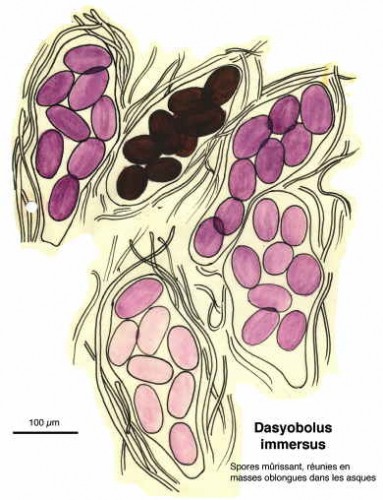
Dessins de André Guyard (1959)
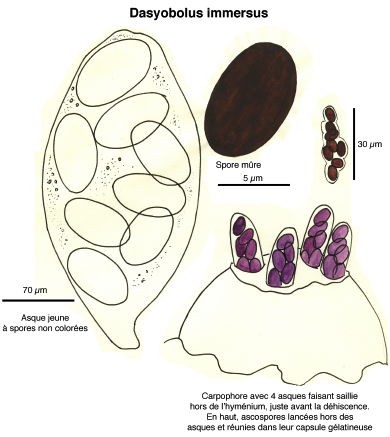
Dessins de André Guyard (1959)
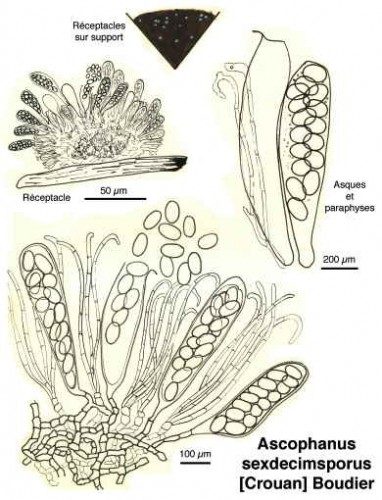
Dessins de André Guyard (1959)
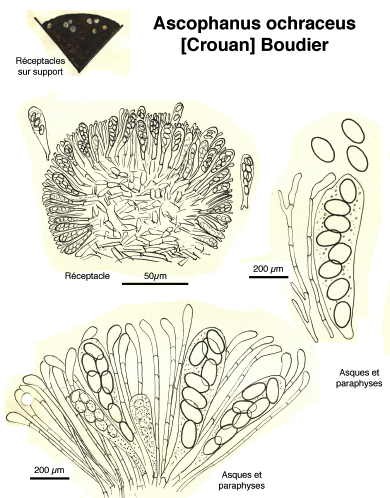
Dessins de André Guyard (1959)
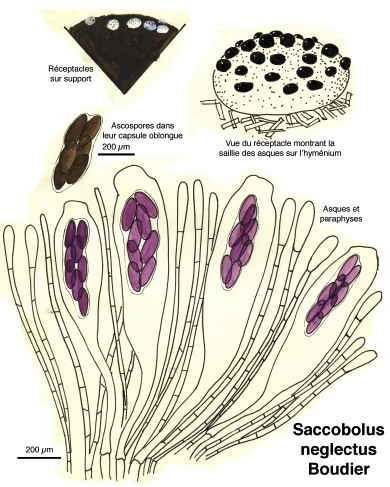
Saccobolus neglectus
Dessins de André Guyard (1959)
Dessins de André Guyard (1959)

Dessins de André Guyard (1959)
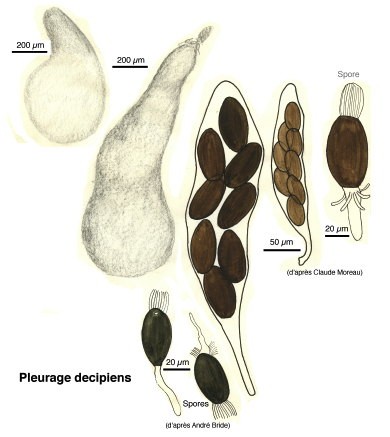
Pleurage decipiens (1)
Dessins de André Guyard (1959)
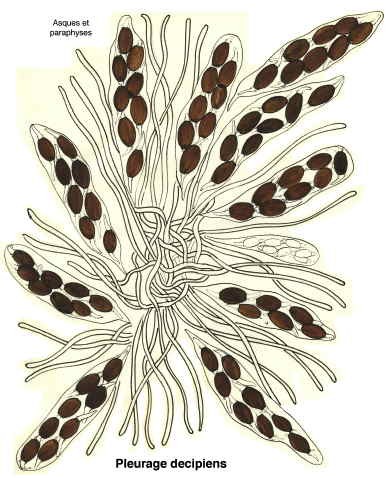
Dessins de André Guyard (1959)
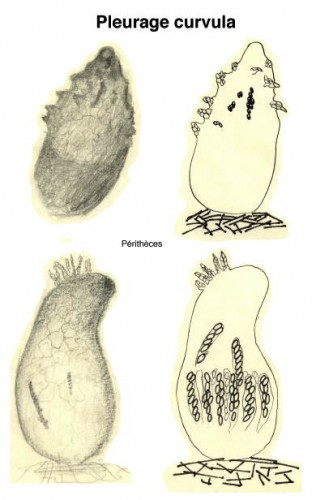
Dessins de André Guyard (1959)
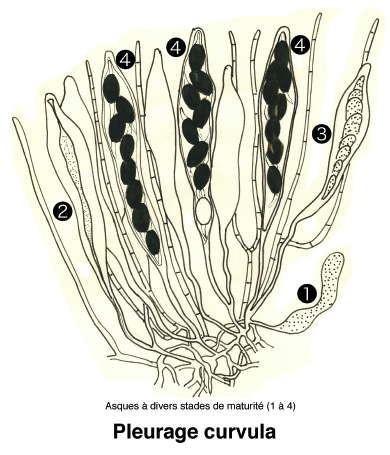
Dessins de André Guyard (1959)
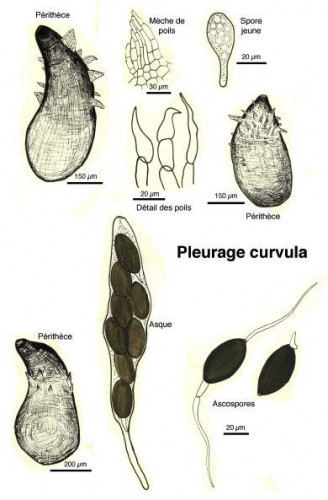
Dessins de André Guyard (1959)
08:22 Publié dans Mycologie | Lien permanent | Commentaires (3) | Facebook | | |
06/10/2009
La Sarcelle Hottentote
La Sarcelle Hottentote
(Anas hottentota) famille des Anatidés
par André Guyard
Ce petit canard fréquente les marais, les lacs et les étangs riches en végétation dans les milieux ouverts. La sarcelle hottentote occupe un vaste territoire qui couvre l'est de l'Afrique et Madagascar.


Pour en savoir plus : http://www.oiseaux.net/oiseaux/sarcelle.hottentote.html
23:58 Publié dans Ornithologie | Lien permanent | Commentaires (0) | Facebook | | |
De Toumaï à Sapiens
 De Toumaï à Sapiens
De Toumaï à Sapiens
La ruée vers l’Homme
par André Guyard
En cette année Darwin, "Quoi de neuf chez nos ancêtres ?"
C’est la question que posent simultanément la Cité des Sciences de Paris ainsi que 14 autres lieux en France dans le cadre d’une même exposition.
Cette expo-dossier – en partenariat avec le Muséum national d’histoire naturelle et le réseau des muséums en région – part à la recherche des origines de l’Homme.
Ces dernières années, de nombreux fossiles d’hominidés ont été mis au jour, qui complètent voire redessinent le scénario de nos origines. Dans le milieu des paléoanthropologues, les polémiques sont parfois vives, le titre de plus vieil ancêtre étant très convoité !
Du 12 février au 7 juin 2009 dans des muséums de province dont celui de Besançon et le musée Cuvier de Montbéliard.
Renseignements : cite-sciences.fr/science-actualites (info : Sciences et Avenir, mars 2009).
Logo de l’article : Reconstitution : Atelier Daynès, Paris © 2003 Ph. Plailly / Eurelios
20:00 Publié dans Préhistoire | Lien permanent | Commentaires (0) | Facebook | | |
Le Tadorne de Belon
 Le Tadorne de Belon
Le Tadorne de Belon
(Tadorna tadorna)
famille des Anatidés
par André Guyard
(dernière mis à jour le 19/02/2016)
Durant tout le mois de janvier 2009, un couple de tadornes de Belon s’est cantonné sur le cours du Trébignon, un exutoire de la nappe phréatique exploitée par la Ville de Besançon par un puits de pompage dit de Chailluz.
Le Trébignon naît à Thise au niveau de la voie ferrée, passe à l’extrémité de l’aérodrome à proximité de champs de maïs et gagne le Doubs au niveau de la piscine de Chalezeule. Le couple a été observé par le garde champêtre de la commune de Thise, des groupes de promeneurs et par Jacques Bonet, le trésorier de la SHND.
Très farouche, à chaque tentative d’approche, les canards prenaient leur envol en direction du Doubs. Étonnant de rencontrer ces volatiles loin du littoral marin où ils fréquentent surtout les côtes marines plates, sablonneuses ou vaseuses. On peut également les observer au bord des lacs salés proches des embouchures ou des estuaires.

Photo de Jacques Nicolin
Le Tadorne de Belon est le plus grand des canards de surface en France. Son plumage bariolé noir, blanc et roux le rend aisément identifiable et le mâle et la femelle sont sensiblement identiques.

D’après la littérature, l’espèce effectue au cours du mois de juillet une migration de mue regroupant adultes nicheurs et non-reproducteurs. Ces rassemblements réunissent plusieurs dizaines de milliers d’individus qui, une fois la mue terminée, regagnent leurs pays d’origine.
Pour assurer sa subsistance à Thise, le couple de tadornes a dû exploiter les champs de maïs.
www.oiseaux.net/oiseaux/tadorne.de.belon.html
Peterson R., Montfort G. & Hollom P.A.D : Guide des oiseaux d’Europe, Delachaux Niestlé Ed.
19:47 Publié dans Ornithologie | Lien permanent | Commentaires (0) | Facebook | | |
Gestion des eaux souterraines
 Gestion des eaux souterraines
Gestion des eaux souterraines
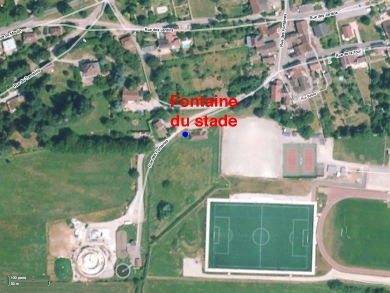
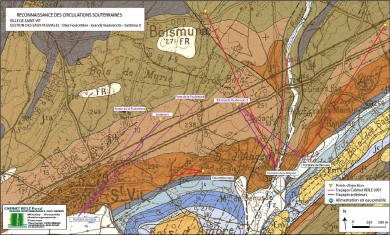
L’exemple de la fontaine du Lavoir ou fontaine du Stade de Saint-Vit (Doubs)
par Pascal Reilé
1 - Problématique
Dans l’optique d’une meilleure connaissance et d’une meilleure gestion des eaux souterraines, la commune de Saint-Vit a fait appel au Comité départemental de spéléologie du Doubs pour explorer l’alimentation de la fontaine du Lavoir (fontaine du Stade) et, en particulier découvrir un prolongement de galerie dans la zone actuellement noyée. Afin de faire la vidange de cette galerie, il faudra procéder à un pompage.

Document : cabinet Pascal Reilé

Document : cabinet Pascal Reilé
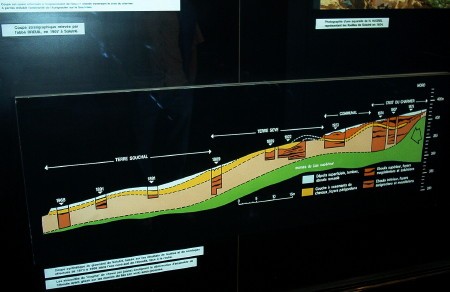
2 – Contexte géologique
La commune de Saint-Vit se trouve à 20 km à l’ouest de Besançon, 20 km au sud de Marnay et 25 km à l’est de Dole. La région est marquée par un grand plissement orienté NE/SO.
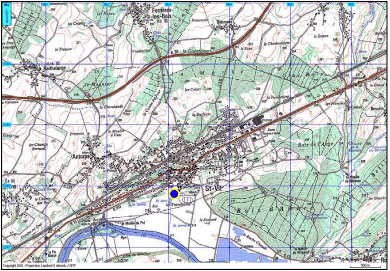
Saint-Vit se situe sur le plateau calcaire encaissant de la plaine alluviale du Doubs, à une altitude moyenne entre 230 et 250 mètres. Ce compartiment présente un faible pendage en direction du Doubs.
La commune se trouve dans le prolongement sud du faisceau bisontin, plissements auxquels appartiennent les anticlinaux de Routelle et d’Osselle. Le compartiment saint-vitois est limité à l’est par un horst qui limite le ruisseau de Sobant et par un anticlinal marqué au niveau de la commune de Routelle.
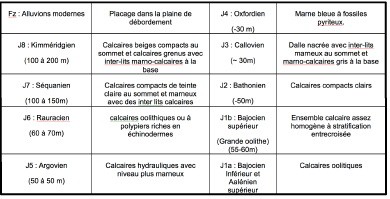
Le sous-sol de la commune est formé de niveaux calcaires du Bathonien (J2). Ces niveaux sont constitués de calcaires beiges massifs à pâte fine et graveleux.
La source qui fait l’objet du pompage se développe dans les formations du bajocien supérieur et inférieur (j1b et jla).
3 – Contexte climatique
Le climat de la région subit une double influence, à la fois océanique et continentale. Le climat général est tempéré humide avec des pluies reparties sur toute l’année, mais le caractère continental s’exprime par des pluies d’été à caractère orageux et des contrastes thermiques de grande amplitude.
Les roches soumises à ce régime climatique associé à une tectonique cassante sont à la fois fissurées et exposées à la dissolution par les eaux chargées en gaz carbonique. La valeur moyenne des précipitations à Besançon/Saint-Vit est de 1 088 mm. La répartition est homogène sur l’ensemble de l’année.
4 – Contexte hydrogéologique
Au niveau de Saint-Vit, il existe deux types de terrains aquifères :
o les alluvions du Doubs, exploitées par la commune de Saint-Vit et par le syndicat du Val de l’Ognon.
o les calcaires karstifiés qui constituent le substratum de la vallée du Doubs (sous les formations alluviales), ainsi que les plateaux de part et d’autre.
Une analyse hydrogéologique sur la connaissance actuelle des écoulements souterrains du secteur de Saint-Vit a permis de réaliser un premier bilan du fonctionnement hydrogéologique pour qualifier le secteur de la commune de Saint-Vit (Cabinet REILÉ - avril 2007).
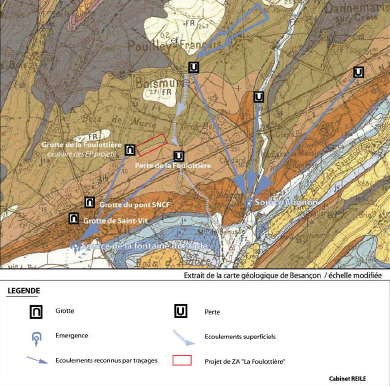
La dynamique des écoulements du secteur Pouilley-Français/Saint-Vit est étroitement liée à un paléo-drainage combinant les écoulements souterrains (karst) et l’écoulement superficiel.
L’affaiblissement des volumes transitant dans les ruisseaux du Rompeux et du Pontot a induit un enfouissement progressif de leurs écoulements dans les calcaires sous-jacents du Jurassique moyen (karstification).
On note une évolution positive de la dynamique du karst dans l’axe SO/NE. Autrement dit : le secteur le plus fossilisé se situe au sud-ouest et le plus fonctionnel au nord-est.
La capture des écoulements souterrains se fait au profit d’un axe renforcé de la perte de Pouilley-Français à la source de la Mignon.
Le secteur ouest de Saint-Vit est fossilisé, la partie est du karst est en cours de fossilisation. La zone de la Foulottière représente une zone à faible fonctionnalité. Le rapprochement des écoulements souterrains du grand drain superficiel est notable.

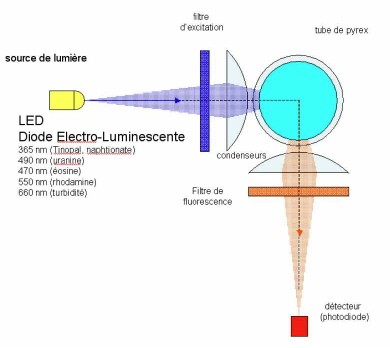
Injection de 3 kg de fluorescéine (Document : cabinet Pascal Reilé)
Document : cabinet Pascal Reilé
Un ensemble de colorations a permis de préciser le fonctionnement des écoulements. Le traçage est quantifié grâce à l’utilisation d’un spectro-fluorimètre comportant une sonde munie d’ une triple optique pour la détection simultanée de 3 traceurs différents alors qu’une quatrième optique mesure uniquement la turbidité de l’eau (voir pour le fonctionnement, l’article intitulé : "Lorsque les eaux du château de Joux ressurgissent à la source de la Loue").
Le site est soumis à des inondations récurrentes dans le lit majeur du Doubs. L’exutoire pompé est en lien direct avec la nappe.

débit mesuré : 1,5 m3/sec (Document : cabinet Pascal Reilé)

débit mesuré : 1,5 m3/sec (Document : cabinet Pascal Reilé)

débit mesuré : 1,5 m3/sec (Document : cabinet Pascal Reilé)
6 – Autres explorations entreprises sur le système du Lavoir
6.1. Sondages de reconnaissance
Le 28 septembre 2007, le cabinet B3G2 a réalisé 23 sondages de reconnaissance dans la doline du terrain de jeux de la commune de Saint-Vit. Les sondages ont permis de définir la dalle calcaire, mais aucun vide karstique n’a été reconnu. Les remplissages par les argiles de décalcification sont majeurs. Le karst est non fonctionnel. Les sondages ont été nivelés topographiquement par le cabinet ROBERT.
6.2 – Dégagement d’un karst perché en bordure de doline – Exploration en cours
Un karst fossile non colmaté existe sur le plateau calcaire qui domine la doline du terrain de jeux de 4 à 5 mètres.

(Document : cabinet Pascal Reilé)

(Document : cabinet Pascal Reilé)
Le dégagement complet du remplissage de la grotte devra être réalisé afin d’éviter tout risque de déstabilisation après avis d’un géotechnicien validant la stabilité des dégagements et aménagements.

Recherche d’exutoire. Dégagement de la grotte et essais d’infiltration (2 et 9 octobre 2008 (Document : cabinet Pascal Reilé)
Remerciements au Cabinet Pascal Reilé qui nous a aimablement communiqué texte et documents écrits concernant cet article.
19:17 Publié dans Géologie-hydrogéologie-Climatologie | Lien permanent | Commentaires (0) | Facebook | | |
Les sauropodes : des géants agiles ?
 Les sauropodes : des géants agiles ?
Les sauropodes : des géants agiles ?
Malgré leur taille gigantesque, les plus grands de tous les animaux ayant existé sur terre étaient capables d’une grande motilité.
par André Guyard
(Mise à jour 05/03/2013)
Parmi les sites dont l’exploitation a donné lieu à l’exposition itinérante Paléomania qui a circulé en 2008 dans l’arc jurassien, nous avons visité Loulle et Coisia dans le Jura français (voir articles consacrés à chacun de ces sites). Il restait à découvrir Courtedoux. [1]
Or les inventeurs du site de Courtedoux, près de Porrentruy dans le Jura suisse, sont à l’initiative de Paléomania et ils ont publié un article dans la revue "Pour la Science" de décembre 2008 concernant les sauropodes de l’arc jurassien.
S’inspirant largement de cet article, la présente note complète les données recueillies lors des visites des sites de pistes de sauropodes de Loulle et de Coisia (Jura français) et montre que, malgré leur taille gigantesque, les sauropodes étaient capables d’une grande motilité.
Le cliché ci-dessous montre des traces de dinosaures imprimé sur le rivage d'une plaine côtière insulaire, il y a 152 millions d’années, à Courtedoux (Jura suisse). La plupart de ces traces rondes (pattes arrière) ou en demi-lune (pattes avant),signent le passage de sauropodes de type Diplodocus,se déplaçant à la recherche de nourriture. Ils étaient suivis de près par de petits théropodes de type Compsognathus (à gauche) et par un grand théropode type Allosaurus, à l’affût peut-être d’un bébé sauropode isolé.


Les sauropodes ont été les plus grands animaux terrestres de l’histoire de la vie.
Argentinosaurus, titanosaure sud-américain du Crétacé supérieur est le plus massif animal terrestre connu. Il mesurait près de 30 mètres de long et pesait près de 90 tonnes. Brachiosaurus, un dinosaure du Jurassique supérieur, qui vivait en Amérique du Nord et en Afrique, observait le monde du haut de ses 12 mètres.

Une fois ces premières étapes complétées, ils ont appliqué leur modèle mathématique au plus grand squelette de dinosaure au monde, le Brachiosaurus brancai du Musée d'histoire naturelle de Berlin. C'est cette étude qui leur a permis de conclure à un poids bien "moindre" que ce qui avait été évalué jusqu'ici. Cette nouvelle méthode est complètement objective, a assuré à l'AFP Bill Sellers soulignant que la masse corporelle est un paramètre très important pour les biologistes. Le pois du Brachiosaurus brancai a été déterminé par les scientifiques à 23 tonnes, alors qu'il était estimé à 80 tonnes depuis les années 1960. Le détail des travaux complets a été publié dans les Biology Letters de la Royal Society.

Brachiosaurus brancai du Musée de Berlin
« Notre méthode fournit une mesure beaucoup plus précise et montre que les dinosaures sont moins massifs qu'on ne le pensait » a déclaré Bill Sellers à The Telegraph. « L’une des choses les plus importantes pour les paléobiologistes est de connaître la masse des animaux fossilisés. C’est étonnamment difficile » ajoute-t-il.
Mais le brachiosaure n'est pas le seul dinosaure qui pourrait voir son poids diminuer avec cette nouvelle méthode "robuste". "Nos résultats suggèrent que de nombreuses estimations précédentes (pour tous les dinosaures) sont vraiment trop lourdes", a ainsi déclaré le Pr Sellers. Pour de nombreux dinosaures, l'écart ne serait pas aussi important que pour le brachiosaure, mais ce sont vraisemblablement "les estimations les plus légères" qui sont correctes, a t-il souligné.
Heinrich Mallison, paléontologue au Musée de Berlin, a salué ces travaux qui sont selon lui révolutionnaires : c'est une « excellente approche » a t-il déclaré. « Non pas l’estimation des tissus mous, mais d’avoir trouvé combien un modèle osseux peut sous-estimer la masse de l'animal tout entier ».
Comment des géants à petit crâne, au long cou, aux membres massifs et à longue queue, embarrassés par leur poids pouvaient-ils être terrestres ? On les supposait même obligés, la plupart du temps, de vivre dans l’eau pour le supporter.
Europasaurus holgeri est un sauropode « nain » de six mètres
Des géants terrestres
Sur terre, les plus grands mammifères sont l’éléphant (cinq à sept tonnes) et le rhinocéros blanc (2,5 tonnes). Un Argentinosaurus pesait donc une quinzaine d’éléphants et quelque 35 rhinocéros.
Les découvertes des plus anciens spécimens en Thaïlande par Eric Buffetaut du CNRS et ses collègues montrent que le gigantisme, véritable « marque de fabrique » des sauropodes, s’est mis en place très tôt dans l’histoire du groupe.
Ces colosses étaient-ils condamnés à vivre dans l’eau, incapables de se déplacer sur la terre ferme sans être écrasés sous leur poids ? Un faisceau d’indices montre au contraire qu’ils étaient bien adaptés au milieu terrestre. Il a fallu attendre les années 1970 et les travaux du paléontologue américain Robert Bakker pour que la biologie comparée révèle leurs aptitudes réelles.

Manifestement aptes à exploiter les niches écologiques des mégaherbivores de leur temps, les sauropodes étaient construits pour s’y déplacer activement, et non pour s’y traîner péniblement. Leurs énormes pattes verticales soutenaient leur corps nettement au-dessus du sol. Robustes et larges, leurs humérus et fémurs pouvaient supporter des efforts autrement plus importants que le simple poids de leur propriétaire.
C’est donc plutôt à la locomotion efficace des éléphants qu’à celle, difficile, des crocodiles à terre qu’il faut comparer la locomotion des sauropodes. Comme ces anciens herbivores géants, les éléphants ont des membres verticaux, qui supportent un poids de plusieurs tonnes. Cela n’empêche nullement les éléphants de se déplacer assez vite (25 kilomètres par heure), même s’ils ne galopent pas, et de se dresser sur leurs pattes arrière pour atteindre un rameau tendre ou pour s’accoupler... Les sauropodes faisaient-ils de même ?
Les nombreuses pistes de sauropodes découvertes sont des indices précieux sur leur locomotion. Au sein d’un même gisement du Jurassique moyen de l’Oxfordshire, en Grande-Bretagne, on a relevé des pistes à voie étroite où les traces de « pieds » et de « mains » sont proches de l’axe du corps, et, à l’inverse, des pistes à voie large. Ces pistes révèlent une différence de posture au sein des sauropodes, entre les diplodocidés (famille du Jurassique supérieur d’Amérique du Nord et d’Afrique), par exemple, et des formes plus évoluées comme les titanosaures (un groupe cosmopolite au Crétacé) dont les pattes étaient plus écartées.
Les pistes montrent aussi que les sauropodes ne laissaient pas traîner leur longue queue (comptant parfois plus de 80 vertèbres), ce qui contredit les anciennes reconstitutions. Le célèbre squelette de Diplodocus qui trône depuis 1908 dans la galerie de paléontologie du Muséum national d’histoire naturelle à Paris a par exemple été monté avec la queue par terre...
En fait, l’articulation des vertèbres caudales indique que les sauropodes maintenaient leur queue à l’horizontale, pour contrebalancer le cou. Cette posture érigée et la queue à l’horizontale impliquent une locomotion plus consommatrice d’énergie que celle des crocodiles, lézards et autres reptiles, dont les membres restent en position latérale et qui traînent leur queue. Pourquoi l’adopter, sinon pour rendre possibles agilité, vitesse et endurance ?
Une grande taille constituait-elle une meilleure défense face aux prédateurs ?
Il existe un autre argument en faveur de l’efficacité de la locomotion des sauropodes : ces géants pouvaient progresser sur des sols mous. L’équipe de Jean-Paul Billon-Bruyat a découvert des empreintes de pieds de plus de un mètre de diamètre à Courtedoux, dans un environnement proche de celui des Bahamas, mais daté du Jurassique supérieur.

Quels avantages sélectifs ont-ils favorisé, dans les environnements jurassiques et crétacés, les lignées de sauropodes de grande taille ? Une grande taille constituait-elle une meilleure défense face aux prédateurs, comme chez les éléphants actuels ?
On peut le penser, mais il n’existait pas a priori de prédateurs capables de s’attaquer à un sauropode adulte. En outre, les pistes de sauropodes montrent qu’ils pouvaient se déplacer en troupeaux, composés de différentes classes d’âge. Comme chez les éléphants, ce comportement grégaire devait protéger les plus jeunes contre les carnivores.
Enfin, l’extrémité de la longue queue de certains sauropodes, tel Diplodocus, servait peut-être de fouet.
Plus convaincante est l’affirmation selon laquelle un sauropode gagnait à être géant pour exploiter les ressources végétales des parties hautes des arbres, inaccessibles aux autres dinosaures herbivores (comme les stégosaures et les ornithopodes). Les sauropodes du Jurassique se nourrissaient dans des environnements dominés par des conifères et devaient avoir un régime alimentaire sélectif. Ils favorisaient la consommation de certains conifères (Araucaria), de ginkgoales (tel le Ginkgo), de certaines fougères (telle Angiopteris) et de prêles (Equisetum), dont les valeurs énergétiques étaient supérieures à celles des cycadales (de type Cycas), des fougères arborescentes et d’autres conifères.
Au Crétacé, les sauropodes ont pu encore diversifier leur nourriture avec l’apparition des plantes à fleurs, tels les magnolias, et goûter aux fruits. Si une partie considérable de la biomasse disponible pour les herbivores se trouvait en hauteur, la grande taille des sauropodes était un avantage crucial.
Comme c’est le cas à Loulle, de très nombreuses pistes de sauropodes ont été retrouvées à proximité d’anciens rivages marins. Se nourrissaient-ils de plantes situées près du rivage, voire d’algues, qui poussaient sur les parties plus élevées des terres émergées ? La question reste ouverte, mais la présence de très grands sauropodes dans ces milieux côtiers montre que les ressources végétales y étaient plus importantes que ne le suggèrent les rares fossiles de flore qui y ont été mis au jour. Pour se sustenter, un sauropode devait sélectionner, ingurgiter et broyer une quantité considérable de végétaux. Un éléphant passe l’essentiel de son temps à chercher sa nourriture ; chaque jour, il consomme environ 200 kilogrammes de végétaux, soit quatre pour cent de son poids, et boit environ 100 litres d’eau. Ces végétaux transitent le long de 40 mètres d’intestins où seulement la moitié de la nourriture est assimilée, un système bien moins efficace que celui des ruminants actuels (telle la vache).
Des pierres pour digérer
Les mâchoires des sauropodes leur servaient avant tout à saisir et à arracher des branchages et des feuillages. Leurs dents pouvaient être spatulées (en forme de cuiller), comme chez les camarasauridés et les brachiosauridés ou encore cylindriques, comme chez les diplodocidés et les titanosaures, mais il est évident qu’elles ne servaient pas à mâcher, contrairement aux dents des grands mammifères herbivores.
Comment digéraient-ils ? Les tissus mous n’étant pratiquement jamais fossilisés, nous ne connaissons pas le système digestif des sauropodes. Toutefois, des gastrolithes (pierres polies) trouvées au sein de la cage thoracique de certains squelettes (240 sur un Seismosaurus de la fin du Jurassique), suggèrent que les sauropodes avalaient des pierres pour faciliter leur digestion comme les pierres de gésier des oiseaux herbivores et granivores actuels.

Ainsi broyés, les végétaux fermentaient à l’intérieur de très longs intestins, à l’aide d’un cocktail de bactéries et de sucs gastriques. Le bol alimentaire fermentait donc lentement dans l’abdomen des sauropodes, dégageant de la chaleur et une grande quantité de gaz intestinaux. Or la fermentation est favorisée par la chaleur, et plus la masse qui fermente est importante, plus la chaleur produite est intense...
À ce propos, le grand corps massif des sauropodes les faisait bénéficier de l’homéothermie de masse, une déperdition de chaleur moins importante due au faible rapport entre la surface du corps et son volume. Pour le diplodocidé Apatosaurus (jusqu’à 30 tonnes), on a pu montrer que la température passe de 25 °C pour un bébé de 12 kilogrammes à environ 41°C pour un adulte de 13 000 kilogrammes.
Avec leur aptitude à conserver la chaleur corporelle, les sauropodes devaient être insensibles aux variations de la température journalière, voire en surchauffe. Ainsi se dégage l’impression que, pour exploiter au mieux les ressources végétales de leur environnement, les sauropodes sont devenus de grandes « cuves à fermentation » sur pattes, alimentées en permanence par un « robot maniable » (leur long cou) portant un "outil de découpe" (leurs mâchoires aux dents coupantes), un système qui gagnait en efficacité en grossissant.
Les tailles gigantesques des sauropodes posent des problèmes biologiques et biomécaniques ardus.
Quel était leur mode de croissance ? Comment supportaient-ils leur poids ? Il apparaît que les dinosaures et les ptérosaures (leurs cousins volants) formaient principalement de l’os de type "fibrolamellaire" le type de tissu osseux que développent les mammifères et les oiseaux, reflet d’une vitesse de croissance élevée. Comme le montre la coupe ci-dessous, le tissu osseux est très riche en canaux vasculaires et présente des cernesde croissance (flèches), indiquant une croissance rapide.
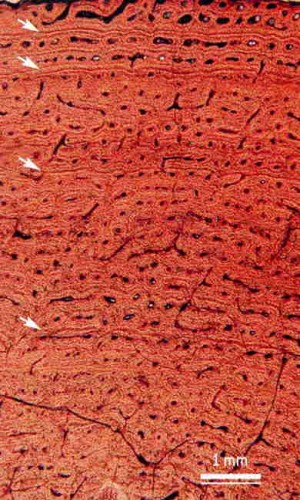
Le sauropode Apatosaurus aurait atteint sa taille adulte en huit à dix ans, ce qui correspond à une prise de masse d’environ 5,5 tonnes par an ! Toutefois, une croissance et une prise de poids rapides posent le problème de la résistance des os, qui soutiennent le corps. Le volume et donc son poids d’un corps en croissance augmentent comme le cube de sa taille. En théorie, le poids d’un sauropode atteignant cinq mètres de haut était 1000 fois plus important que celui du même animal quand il ne mesurait que 50 centimètres. Et chaque centimètre carré de section du fémur d’un sauropode de cinq mètres de haut devait supporter dix fois plus de pression qu’un centimètre carré du même fémur quand l’animal mesurait seulement 0,5 mètre de haut.
Le squelette des sauropodes, bien ancré sur des membres massifs et des ceintures scapulaires (les épaules) et pelvienne (le bassin) robustes, présentait-il des allégements ? Un exemple est donné par leur long cou, constitué de vertèbres pourvues d’un système complexe de cavités. Dès les premières découvertes de squelettes de sauropodes, les paléontologues ont pensé qu’elles avaient évolué pour alléger la longue colonne vertébrale. Bien que les vertèbres des sauropodes soient massives, leurs structures sont comparables à celles des oiseaux en ceci qu’elles sont pleines d’air en connexion avec les sacs aériens et les poumons.
Pour autant, l’allégement de la colonne vertébrale et ce système respiratoire de type avien étaient-ils propres aux sauropodes ? Non, on les rencontre aussi chez les prosauropodes (dinosaures herbivores du Trias et du Jurassique inférieur, apparentés aux sauropodes), les théropodes et les ptérosaures ; mais ils sont absents chez les autres reptiles.
Le long cou des sauropodes
Le cou des sauropodes avait donc des vertèbres allégées, mais il était aussi démesuré, si long qu’il en paraît aberrant, jusqu’à quatre fois la longueur du tronc.
Comment les sauropodes ont-ils développé de si longs cous ? Comment la circulation du sang contrait les effets de la gravité pour irriguer le cerveau ? Ce cou était-il flexible ?
Nous avons vu que ce long cou leur était nécessaire pour « ratisser » efficacement leur environnement. Étant donné la longueur, la posture et la souplesse de leurs cous, on déduit les caractéristiques principales des niches écologiques qu’occupaient les sauropodes. On a pu démontrer que le cou de Brachiosaurus aurait été quasiment à l’horizontale. La grande taille de Brachiosaurus et ses membres antérieurs, plus longs que les postérieurs, lui permettaient de saisir des branches situées à plus de six mètres de haut. L’articulation des vertèbres cervicales de Diplodocus montre que son très long cou était peu flexible vers le haut ; en revanche, il pouvait aisément le baisser pour manger à même le sol. Ainsi, Diplodocus pouvait saisir des feuillages à quatre mètres de haut, mais aussi brouter des fougères et des prêles, avec l’élégance d’un bovin.
Le cou des sauropodes s’est allongé au fil du temps en empruntant deux voies : un allongement de la longueur des vertèbres cervicales (atteignant parfois plus d’un mètre) ou une augmentation de leur nombre (jusqu’à 19) par conversion de vertèbres dorsales en cervicales, voire les deux simultanément. Le très long cou de Brachiosaurus, par exemple, résulte d’un allongement de ses cervicales (13 vertèbres dont certaines atteignent 75 centimètres).
À l’extrémité du cou des sauropodes se trouvait un crâne minuscule, en proportion avec le corps : 70 centimètres de long pour un Diplodocus de 26 mètres ! Les mâchoires des sauropodes ne servant pas à mastiquer, mais seulement à la prise de nourriture, leur crâne a pu rester petit, prérequis nécessaire au développement d’un long cou. De fait, les sauropodes avaient le plus faible quotient d’encéphalisation de tous les dinosaures, c’est-à-dire le plus faible rapport masse cérébrale/masse corporelle.
Le quotient d’encéphalisation et l’anatomie des sauropodes posent la question de l’irrigation d’un cerveau situé à l’extrémité d’un si long cou. Afin de contrer les effets de la gravité étant donné leur anatomie, les sauropodes ont dû, comme la girafe, développer une pompe cardiaque puissante et des vaisseaux sanguins renforcés et équipés de valves de sécurité. La pression artérielle de Brachiosaurus, dont la tête s’élevait jusqu’à 12 mètres au-dessus du sol et à huit mètres au-dessus de son cœur, devait ainsi être très élevée. On a estimé la différence de pression entre sa tête et son cœur à 600 millimètres de mercure !
Des quadrupèdes herbivores gigantesques plus proches des oiseaux que des reptiles
Les puissants muscles nécessaires pour animer un énorme corps suggèrent que le métabolisme des sauropodes était élevé, pareil à celui des animaux homéothermes (à température constante) et endothermes (production de chaleur interne), que l’on rencontre chez les mammifères et les oiseaux. Une étude géochimique a pu établir que les dinosaures avaient une physiologie thermique plus proche de celle des mammifères que de celle des reptiles contemporains.
Ainsi, les plus grands vertébrés terrestres ayant jamais vécu, les sauropodes, étaient des quadrupèdes herbivores dynamiques, construits pour optimiser leur quête de nourriture et leur digestion. Leur biologie, caractérisée par un métabolisme élevé, était bien plus proche de celle des oiseaux et des mammifères que de celle des autres reptiles. Ce résultat est conforté par la classification des espèces puisque, sur la branche des dinosaures, les sauropodes sont proches des théropodes, dont descendent les oiseaux.

Succès évolutif des Sauropodes
(Pour la Science mars 2013)
Pour zoomer, cliquer sur le cliché
• BIBLIOGRAPHIE
J.-P. Billon-Bruyat, D. Marty et D. Becker, Les sauropodes, géants agiles, Pour la Science, n° 374, décembre 2008.
J. Horner, K. Padian et A. de Ricqlès, Dinosaures : les secrets de leur taille, Pour la Science, n° 334, août 2005.
J. Hummel et ., In vitro digestibility of fern and gymnosperm foliage : implications for sauropod feeding ecology and diet sélection, Proceedings of the Royal Society B, vol.275, pp. 1015-1021, 2008.
D. Marty et al., Late Jurassic dinosaur tracksites of the Transjurane highway (Canton Jura, Switzerland) : overview and measures for their protection and valorisation, Bulletin for Applied Geology, vol. 12, pp. 75-89. 2007.
D. Schwarz et G. Fritsch, Pneumatic structures in the cervical vertebrae of the Late Jurassic Tendaguru sauropods Brachiosaurus brancai and Dicraeosaurus , Ecologae Geologicae Helvetiae, vol. 99, pp.65-78, 2006.
A. Bénéteau : http://www.paleospot.com
http://www.lejurassique.com/lejurassique/fr/paleomania.html
Vifs remerciements à Jean-Paul BILLON-BRUYAT, Daniel MARTY et Damien BECKER qui m’ont autorisé à reproduire de longs passages du texte de leur article dans "Pour la Science" de décembre 2008.
Jean-Paul BILLON-BRUYAT, Daniel MARTY et Damien BECKER sont paléontologues à la Section d’archéologie et paléontologie du Canton du Jura, en Suisse.
17:34 Publié dans Paléontologie - archéologie | Tags : dinosaures, sauropodes, fossiles, pistes, jurassique | Lien permanent | Commentaires (2) | Facebook | | |
Le retour du Saumon dans le Rhin suisse
 Le retour du Saumon dans le Rhin suisse
Le retour du Saumon dans le Rhin suisse
par André Guyard
Un saumon femelle a été observé dans le Rhin à Bâle le dimanche 5 octobre 2008. Ce saumon a été formellement identifié par des experts. Bien sûr, il a été soigneusement relâché car la pêche d’un saumon ou d’une truite de mer est interdite dans le bassin du Rhin.

Il est forcément issu d’un alevinage des autorités piscicoles suisses, mais nul ne sait comment il a réussi à remonter le cours si haut en l’absence de dispositifs techniques pour assurer la continuité écologique du fleuve au-delà de Gambsheim. Il a dû emprunter des écluses de navigation, on n’a pas fini de s’interroger, mais c’est quasi miraculeux.


- faire construire une passe à poissons à Strasbourg,
- faire construire une passe à Gerstheim et sur les ouvrages dits "barrages agricoles" des festons de Strasbourg, Gerstheim et Rhinau. Ces travaux doivent permettre l’accès du saumon à Fribourg avant 2015. Aujourd’hui un an après la décision, EDF n’a strictement rien fait et utilise des mesures dilatoires pour essayer d’échapper à l’obligation de financer tout seul les ouvrages.

En outre, un courrier de la France aux autorités allemandes pour leur demander d’apporter une contribution financière serait resté à ce jour (octobre 2008) sans réponse.
17:09 Publié dans Environnement-Écologie | Lien permanent | Commentaires (0) | Facebook | | |
La Poule d’eau
 La Poule d’eau
La Poule d’eau
Gallinula chloropus (Famille des Rallidés)
par André Guyard
On confond souvent la Poule d’eau ou Gallinule avec la Foulque macroule (Fulica atra). La Gallinule s’en distingue par sa taille inférieure et surtout par sa plaque frontale rouge et son bec rouge terminé par une pointe jaune (plaque frontale et bec blancs chez la Foulque).


La Poule d’eau vit près des eaux douces dans des zones humides où la végétation est abondante et émergente proposant un couvert. C’est un très bon plongeur, restant parfois plus de 45 secondes sous l’eau. Elle nage ou marche le long des rives, ou court se mettre à couvert.

On la rencontrera de préférence dans les petits cours d’eau et les marécages.



Les poussins sont nidifuges. Ils sont souvent couvés pendant les deux premières semaines, surtout par temps froid et humide. Les parents et les jeunes des premières portées s’occupent des poussins pendant parfois 6 semaines. Les jeunes sont indépendants vers 3 ou 4 semaines.
16:30 Publié dans Ornithologie | Lien permanent | Commentaires (0) | Facebook | | |
Le Phalarope à bec étroit
 Le Phalarope à bec étroit
Le Phalarope à bec étroit
Phalaropus lobatus (Scolopacidés)
par André Guyard
Les Phalaropes sont de gracieux limicoles, d’une taille d’une quinzaine de cm, nageant avec vivacité souvent en virant sur eux-mêmes en eau peu profonde, créant ainsi un tourbillon pour amener à la surface le plancton dont ils se nourrissent.
L’espèce niche en groupes dans les zones humides, les îles des rivières et les plages aux bords des lacs dans le nord de l'Europe.
Le Phalarope à bec étroit est une espèce moins nordique que sa cousine à bec large. En Europe, elle niche en Islande, dans les îles écossaises, les chaînes scandinaves et la toundra nordique. En automne, elle entreprend un long trajet de migration. Grâce à un GPS pesant 0,6 g, supportable pour ces limicoles de 20 cm de long vivant l'été en Écosse et en Islande, l'Institut ornithologique suisse a pu déterminer en 2013 que l'oiseau ne migrait pas en Arabie Saoudite l'hiver comme on le pensait, mais en Équateur et au Pérou. Soit plus de 10 000 km de voyage aller.
C’est une espèce rarement observée en France le long des côtes atlantiques.


C'est la femelle qui défend le territoire tandis que le mâle construit le nid, assure seul l’incubation qui dure entre 18 et 20 jours, ainsi que l’éducation des poussins. C'est donc en toute logique, que le plumage de la femelle est plus coloré que celui du mâle.
Il arrive que la femelle ponde une seconde fois, avec un autre partenaire.

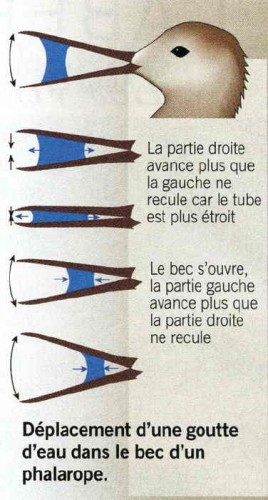
Schéma dû au talent de Sylvie Daoudal
L’extrémité de la goutte qui se trouve proche de la bouche avance alors plus que l’extrémité opposée. En rouvrant le bec, cette fois la partie « gauche » se déplace plus que la « droite ». Après un cycle de claquements de bec, la goutte progresse vers la bouche. Les chercheurs ont observé qu’il existe des angles optimaux permettant de déplacer la goutte en un petit nombre de claquements. Le Phalarope satisfait à cet idéal avec deux coups pour cinq centimètres de déplacement et ce à raison de deux gouttes par seconde. D’autres oiseaux (bécasseaux, échasses) dont ce n’est pas le seul mode d’alimentation, ont besoin de cinq à sept coups. Malgré la gravitation, l’eau ne tombe pas, retenue par les aspérités de la surface du bec (telles les gouttes de pluie sur les vitres verticales).
Une belle mais fragile adaptation : les chercheurs soulignent l’extrême sensibilité de ces espèces à la pollution des eaux. La présence de savon ou d’huile modifierait les conditions de mouillage de l’eau, au point d’empêcher l’ascension des gouttes et de leur contenu dans le bec. L’espèce est également menacée par le drainage des zones humides, l’intensification des habitats, les dérangements humains, les inondations et la prédation, notamment par le Labbe parasite (Stercorarius parasiticus).
Pour en savoir plus : http://www.oiseaux.net/oiseaux/phalarope.a.bec.etroit.html
16:20 Publié dans Ornithologie | Lien permanent | Commentaires (0) | Facebook | | |
Le Plésiosaure de Palente
 Le Plésiosaure de Palente
Le Plésiosaure de Palente
par André Guyard
(Dernière mise à jour : décembre 2014)
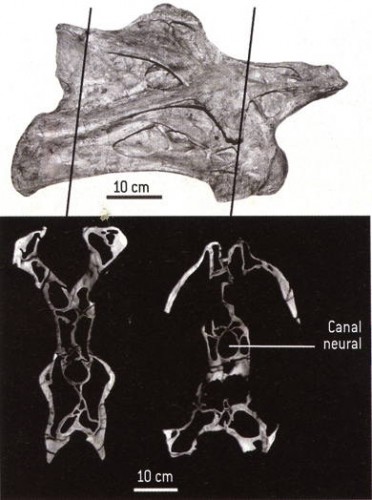
En décembre 2006, à l’occasion des travaux d’aménagement de l’entrée Est de Besançon, le regard averti de Nicolas Martin, étudiant en master de géologie appliquée de l’Université de Franche-Comté a permis la découverte d’ossements de plésiosaure, grand reptile marin du Jurassique.
Avec la complicité de la Communauté d’Agglomération du Grand Besançon et des entreprises du chantier, une fouille de sauvetage a pu être organisée par le département de Géosciences de l’Université. Cette fouille dirigée par Vincent Bichet, maître de conférences en Géosciences, a permis la mise au jour d’une quarantaine de vertèbres appartenant à un plésiosaure.
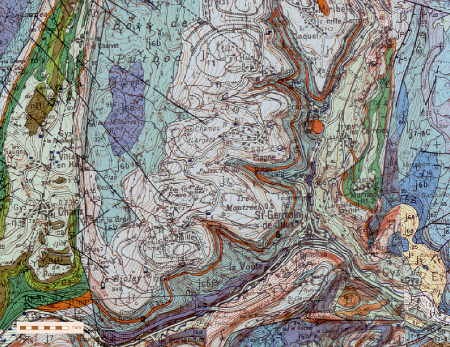
La zone prospectée se situe au niveau du cercle rouge pâle
Le site se localise au pied du Fort Benoît dans le quartier de Palente au lieu-dit les Tuileries, au niveau du 2e giratoire. Le site doit son nom au fait que ces argiles grises datées du Callovo-Oxfordien ont été exploitées dans des carrières pour la fabrication de tuiles et de briques (voir carte géologique).
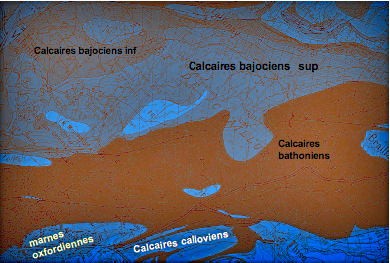
Au sud de la carte se situent les marnes callovio-oxfordiennes (en bleu)
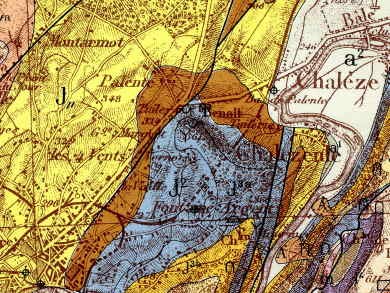
Le lieu-dit "Les Tuileries" se situe sur les marnes callovio-oxfordiennes (en bleu)

Les argiles marneuses du Callovien affleurent au niveau
Du point de vue stratigraphique, les calcaires du Callovien qui surplombent les calcaires du Bathonien sont peu épais (15 à 20 m). Ce sont des calcaires coquilliers et oolithiques, similaires à ceux du Bathonien, qui se débitent en dalles de 3 à 10 cm d’épaisseur et qui ont été employées dans le passé pour la couverture des maisons (et appelées laves dans la région). Ces calcaires constituent le substratum des quartiers des Quatre-vents et des Orchamps et de la place des Tilleuls à Palente.

(Cliché : Vincent Bichet)
La paléogéographie
C’est donc à ce niveau qu’ont été trouvées une quarantaine de vertèbres appartenant à un plésiosaure. Ainsi les eaux marines calloviennes étaient parcourues par ces reptiles marins carnassiers qui chassaient la faune pélagique comme les ammonites et les bélemnites. Des îles émergeaient, couvertes de végétaux, dont les débris ont donné des bois flottés.

(Cliché : Vincent Bichet)

(Cliché : Vincent Bichet)
[2] Faune benthique : animaux vivant sur le fond.
Crédits photos : Vincent Bichet, maître de conférence au département Géosciences de l’Université de Franche-Comté.
Remerciements à Patrick Rolin, maître de conférence au département Géosciences de l’Université de Franche-Comté qui nous a fourni des éléments pour construire le texte.
C'est désormais chose faite avec la découverte, dans l'est de la Chine (province d'Anhui), des restes fossilisés d'un ichthyosaure amphibie vieux de 248 millions d'années. “Cette nouvelle espèce, baptisée Cartorhynchus lenticarpus, a des nageoires anormalement larges et courbées par rapport aux ichtyosaures postérieurs. Elles devaient lui permettre de supporter son poids lorsqu'il se déplaçait sur la terre ferme, comme le font aujourd'hui les phoques” explique Ryosuke Motani. l'un des chercheurs de l'université de Californie à l'origine de cette découverte.
Comme les phoques, Cartorhynchus lenticarpus possédait un museau court. Avec ses 40 cm de long, c'est aussi le plus petit ichtyosaure connu, bien loin de la dizaine de mètres qu'ont atteint d'autres ichtyosaures.

Cartorhynchus lenticarpus © Science & Vie
16:02 Publié dans Paléontologie - archéologie | Tags : dinosaures, jura, jurassique | Lien permanent | Commentaires (0) | Facebook | | |
Halte à l’orpaillage clandestin en Guyane !
La Guyane concentre sur son territoire un grand nombre d'exploitations minières illégales générant une insécurité importante et des atteintes environnementales quasi irrémédiables (déforestation massive et sauvage, propagation de métaux lourds dans les fleuves et le sous-sol). L'orpaillage illégal s'appuie sur des zones logistiques situées pour la plupart à l'extérieur du département, et à partir desquelles les sites d'exploitation sont alimentés en moyens humains et matériels par voie fluviale, puis en empruntant un maillage de pistes et layons tracés par les délinquants.
15:42 Publié dans Environnement-Écologie | Tags : environnement, pollution, guyane, orpaillage | Lien permanent | Commentaires (0) | Facebook | | |
La Mouette rieuse
 La Mouette rieuse
La Mouette rieuse
Larus ridibundus
(Famille des Laridés)
par André Guyard
La Mouette rieuse appartient à la famille des Laridés comme les goélands et les sternes. Chez la Mouette rieuse, les deux sexes sont identiques.

En juillet, la Mouette rieuse se coiffe d’un capuchon chocolat
Pour en savoir plus :
www.oiseaux.net/oiseaux/mouette.rieuse.html
15:33 Publié dans Ornithologie | Lien permanent | Commentaires (0) | Facebook | | |
La Marmotte des Alpes
 La Marmotte des Alpes
La Marmotte des Alpes
Marmota marmota
(Linnæus, 1758)
Famille des Sciuridés
(Dernière mise à jour : 15/08/2018)
par André Guyard
Silhouette trapue, petites oreilles se dissimulant dans une fourrure épaisse, museau large et court, pattes puissantes adaptées au fouissage, la marmotte court ventre à terre dans les alpages pour se réfugier dans son terrier en cas d’alerte.



La Marmotte se nourrit de graminées, légumineuses, graines et baies, racines et fleurs. Mais elle ne dédaigne pas les insectes comme les coléoptères ou les orthoptères.
Pendant le rut qui survient après le réveil, les mâles défendent leur territoire qu’ils marquent avec leurs glandes jugales. La gestation dure 5 semaines. La femelle met au monde une portée de 2 à 4 petits qui sortiront du terrier au bout de 5 semaines. La maturité sexuelle survient après la 3e hibernation.
À partir de l'âge de 2 ans — celui auquel ils quittent leurs parents —, les jeunes marmottons nés hors couples stables ont une chance de survie accrue de 30 %, note Aurélie Cohas. Enfin, les rejetons adultérins ont un succès reproducteur légèrement meilleur et accèdent plus fréquemment à des statuts de dominants.
Son principal prédateur : l'Aigle royal qui s'installe dans les Alpes, mais qui essaie de s'établir dans le Haut-Jura (où il n'y a pas de marmottes !). Voir le reportage de France 3 sur l'Aigle royal.
Dormir comme une marmotte : l’expression a fait florès. En effet, ce rongeur de la famille des Sciuridés (tout comme l’écureuil) échappe aux mauvaises conditions hivernales en se plongeant dans un profond sommeil.
11:37 Publié dans Mammifères | Lien permanent | Commentaires (0) | Facebook | | |
Le Macareux moine
 Le Macareux moine
Le Macareux moine
Fratercula arctica
(Famille des Alcidés)
par André Guyard
Grâce à son bec triangulaire légèrement crochu, à pointe rouge, base bleu foncé cernée de jaune, le Macareux Moine est facile à identifier. Il utilise son bec pour stocker ses proies. Il avale ses proies sous l’eau, sauf quand il nourrit son poussin. Ses ailes courtes le prédisposent à la nage sous l’eau. Pour le vol, leur faible portance contraint l’oiseau à un battement très rapide. L’aire de répartition du Macareux Moine est strictement nord atlantique. C’est un oiseau pélagique qui vit en haute mer. Il se rend sur la terre ferme uniquement en période de reproduction. Il niche alors sur les pentes herbeuses et les falaises côtières d’Islande (2 à 3 millions de couples), d’Irlande, d’Écosse, des Shetland et de Scandinavie... Une population relictuelle subsiste en Bretagne.

Le Macareux niche en colonies sur les falaises


Pour en savoir plus :
11:27 Publié dans Ornithologie | Lien permanent | Commentaires (0) | Facebook | | |
Le lapiaz de Loulle (Jura)

Le lapiaz de Loulle (Jura)
par André Guyard
Non loin de Champagnole (Jura), le village de Loulle présente un lapiaz découvert, creusé dans les calcaires du Séquanien. « Lapiaz » est un terme jurassien issu étymologiquement du latin : lapis (la pierre).
Facilement accessible et spectaculaire, ce phénomène karstique se révèle à découvert au milieu des prairies voisines.

Un lapiaz se présente comme une dalle calcaire dont les fissures se développent en sillons.
Les sillons sont de deux types :
- les rigoles rectilignes ou sinueuses, suivant la ligne de la pente ou non ;
- les crevasses (ou leisines), qui sont un approfondissement des diaclases et qui découpent la roche en blocs.

Pour qu’un lapiaz se développe, il est nécessaire d’avoir une dalle calcaire surmontée d’une couche d’humus dont le lixiviat sera très acide, et un faible pendage des couches permettant un long ruissellement de l’eau au contact du calcaire.
Ainsi, l’eau de pluie acidifiée par le CO2 atmosphérique et les acides humiques s’attaquera au calcaire sous-jacent, formant des rigoles de dissolution.
Le CO2 soustrait de l’atmosphère se transforme en ions HCO3- selon la réaction suivante :
Ca CO3 + CO2 +H2O => 2HCO2- + Ca2+
Le lapiaz de Loulle aujourd’hui mis à nu et non actif, s’est probablement formé sous un sol forestier qui n’apparaît plus aujourd’hui, érodé par les glaciers présents encore il y a 20 000 ans dans le Jura.
Les fissures et cavités du lapiaz constituent des microbiotopes hébergeant toute une flore particulière comme cette langue de bœuf ou Scolopendre, Asplenium scolopendrium,

ou d’autres délicates fougères comme cette Gymnocarpium robertianum.

10:24 Publié dans Géologie-hydrogéologie-Climatologie | Lien permanent | Commentaires (0) | Facebook | | |
Le Lagopède des Alpes
 Le Lagopède des Alpes
Le Lagopède des Alpes
Lagopus mutus
(Famille des Tétraonidés)
par André Guyard
Un couple de Lagopèdes des Alpes a été rencontré en Islande en juillet 2008 le long d’un sentier de randonnée pédestre. Les oiseaux sont alors en plumage d’été. Le mâle se distingue de la femelle par sa caroncule rouge bien visible et par une barre noire qui va de la base du bec à l’arrière de l’œil. En hiver, mâle et femelle présentent un plumage blanc, à l’exception de la queue qui reste noire.


La femelle ne manifeste aucun effroi à l’approche des randonneurs

La femelle en livrée estivale a un plumage qui la camoufle lors de la couvaison
Pour en savoir plus :
http://www.oiseau-libre.net/Oiseaux/Especes/Lagopede-alpi...
10:07 Publié dans Ornithologie | Lien permanent | Commentaires (0) | Facebook | | |
L’Huîtrier pie
 L’Huîtrier pie
L’Huîtrier pie
Haematopus ostralegus
(Famille des Haematopodidés)
par André Guyard
L’Huîtrier pie est un oiseau de rivage. Ce robuste limicole est doté d’un long bec rouge orangé. Son plumage est entièrement noir et blanc, ce qui lui vaut le qualificatif de "pie". Il n’y a pas de dimorphisme sexuel.

L’Huîtrier pie visite l’estran
Les couples nichant en France, surtout en Normandie et en Bretagne, sont sédentaires. En automne, ils sont rejoints par les individus du nord de l’Europe : Islande, Pays-Bas et Îles britanniques. Les hivernants repartiront au printemps.
L’Huîtrier pie niche en couples séparés, mais devient très sociable après la période de nidification. Le nid est constitué d’une simple dépression garnie de coquillages et de débris d’algues. Entre mars et mai, la femelle y dépose en moyenne trois œufs jaunâtres striés de brun qui sont couvés pendant une période d’un mois environ.

Pour en savoir plus :
www.oiseaux.net/oiseaux/huitrier.pie.html
09:53 Publié dans Ornithologie | Lien permanent | Commentaires (0) | Facebook | | |
Quel était, au Néolithique, le rôle des haches en jades alpins ?
 Quel était, au Néolithique, le rôle des haches en jades alpins ?
Quel était, au Néolithique, le rôle des haches en jades alpins ?
Colloque du 24 au 26 septembre 2009 à Besançon



Différents modules de lames de haches en "jade" alpin

Aquarelle de Benoît Clarys (Pour la Science, septembre 2009)
jade.colloque@mshe.univ-fcomte.fr
09:35 Publié dans Préhistoire | Lien permanent | Commentaires (0) | Facebook | | |
La Grive mauvis
 La Grive mauvis
La Grive mauvis
Turdus iliacus
(Famille des Turdidés)
par André Guyard
Les grives appartiennent à la famille des Turdidés qui regroupe grives, merles, traquets, rougequeue et rougegorge. Oiseau d’Europe septentrionale (Islande et Scandinavie), la Grive mauvis est une grive de petite taille qui ressemble à la Grive musicienne. Elle s’en distingue par un sourcil blanc crème très apparent et les flancs et le dessous des ailes roux vif. Pas de dimorphisme sexuel. La Grive mauvis passe l’hiver en Europe occidentale et méditerranéenne.

Sourcils blancs, flancs et dessous des ailes roux caractérisent la Grive mauvis
La Grive mauvis habite l’Europe septentrionale. C’est un oiseau typiquement de la taïga et de la toundra. Elle niche de préférence dans les forêts de conifères, mais en Islande, elle niche dans les bois de bouleaux tortueux. Pas farouche, l’espèce fréquente parcs et jardins.

Mâle et femelle bâtissent leur nid ensemble qu’ils installent dans les arbres ou les arbustes, un nid façonné de terre glaise et garni d’herbe et de feuilles. La femelle couve seule 4 à 6 œufs. L’incubation dure 14 jours. Les oisillons quittent le nid au bout de deux semaines, mais continuent à être nourris par les parents durant un certain temps.
La Grive mauvis possède un régime omnivore. Elle prospecte surtout au sol et dans les buissons à la recherche d’insectes, de vers et de fruits.
09:25 Publié dans Ornithologie | Lien permanent | Commentaires (0) | Facebook | | |
Le Grèbe esclavon
 Le Grèbe esclavon
Le Grèbe esclavon
Podiceps auritus
(Famille des Podicipitidés)
par André Guyard
Paré de son plumage nuptial, le Grèbe esclavon a le cou et les flancs marron roux ; la tête noire s’empanache de deux huppes jaune doré qu’il perd en hiver.

Nicheur en Europe du Nord : Islande, Scandinavie, Pays baltes, il affectionne les nappes d’eau peu profondes avec une végétation émergeante abondante et une part d’eau libre. Le nid, formé de débris végétaux est dissimulé dans les hélophytes. En mai juin, la femelle pond 3 à 6 œufs dont l’incubation dure entre 22 et 25 jours.

Le Grèbe esclavon se nourrit d’insectes aquatiques, de batraciens, de mollusques, de crustacés et de petits poissons ainsi que de plantes flottantes ou rivulaires.
En novembre, les grèbes migrent vers le sud de préférence sur la façade maritime. En France intérieure, on ne l’observe que sur de vastes plans d’eau comme le lac de Der en Champagne ou le lac de Madine en Lorraine.
La vidéo ci-dessous a été réalisée en juillet 2008 dans l'ouest de l'Islande par André Guyard
Pour en savoir plus :
www.oiseaux.net/oiseaux/grebe.esclavon.html
09:18 Publié dans Ornithologie | Lien permanent | Commentaires (0) | Facebook | | |
Le Fulmar boréal
 Le Fulmar boréal
Le Fulmar boréal
Fulmarus glacialis
(Famille des Procellariidés)
par André Guyard
Le Fulmar boréal (précédemment appelé Pétrel fulmar) appartient à la famille des Procellariidés qui regroupe pétrels et puffins. C’est le seul représentant de cette famille qui soit doté d’un plumage clair et d’une vie diurne.
Le Fulmar est un magnifique voilier. Ses ailes rigides, longues et étroites lui permettent de planer et de glisser au ras des vagues. Cet oiseau pélagique évolue uniquement en haute mer sans guère se poser. Il peut boire l’eau de mer et il excrète le sel par ses narines tubulaires. Il se nourrit de poissons, de plancton, de céphalopodes (calmars, seiches), ou de crustacés.

Le Fulmar niche sur les corniches des falaises côtières.

Le poussin est cette petite boule grise qui émerge derrière le pissenlit
Pour en savoir plus :
http://www.oiseaux.net/oiseaux/fulmar.boreal.htm
09:10 Publié dans Ornithologie | Lien permanent | Commentaires (0) | Facebook | | |
La source intermittente de Fontaine-Ronde
 La source intermittente de Fontaine-Ronde
La source intermittente de Fontaine-Ronde
par André Guyard
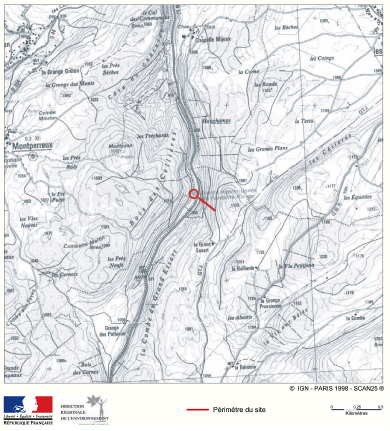
Carte IGN
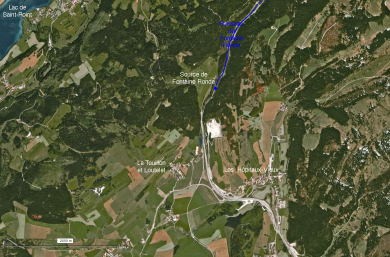
En bleu, cours souligné du ruisseau de Fontaine Ronde
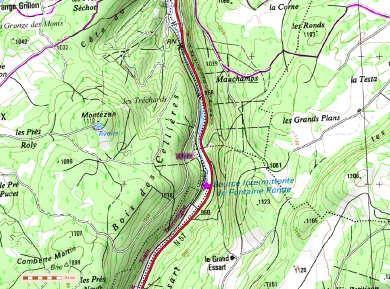
Carte IGN
Des études hydrobiologiques s’appuyant notamment sur la méthode de l’I.B.G.N. (indice biotique global normalisé) mise au point au Laboratoire d’Hydrobiologie de l’UFR Sciences de Franche-Comté ont montré que la qualité du site aquatique était excellente. Le ruisseau de Fontaine Ronde, jusqu’à son confluent avec le Doubs à proximité du Château de Joux constitue l’un des derniers cours d’eau de Franche-Comté à montrer une telle biodiversité, en particulier pour la faune benthique (larves de Plécoptères indicatrices d’une excellente qualité des eaux).
Les analyses chimiques ont montré une teneur élevée en chlorure de sodium (NaCl), à rapprocher de la composition des eaux de la poche phréatique percée lors de la construction du tunnel du Mont d’Or (voir plus loin).

La source se présente sous la forme d’une vasque
Protégée par un bassin maçonné semi-circulaire, la source de Fontaine Ronde se situe en contrebas du remblai de la voie ferrée actuellement désaffectée mais en cours de restauration pour un train touristique, le Coni’fer. Elle se présente sous la forme d’une vasque au fond sableux d’environ 4 m de diamètre. Il s’agit d’une source vauclusienne qui présente un débit intermittent. Cette source intermittente (il n’y a que sept sources intermittentes en France et une trentaine dans le monde) laisse apparaître toutes les 10 minutes environ une faible montée des eaux. L’amorce de cette montée est précédée d’une apparition de bulles de gaz (d’air ou de CO2 ?).
La source a la particularité d’observer un débit dont les variations étaient autrefois régulières dans le temps. De plusieurs dizaines de centimètres jusqu’au début du XXe siècle, le battement est réduit aujourd’hui à quelques centimètres et la périodicité est désormais irrégulière. C’est en 1912, lors du creusement du tunnel ferroviaire du Mont d’Or, distant pourtant de plusieurs kilomètres, qu’une poche de la nappe phréatique alimentant toutes les sources du secteur fut rompue, inondant le tunnel et tarissant les sources qu’elle alimentait : grande source de Malbuisson, source du Bief Rouge à Métabief, dont la Fontaine Ronde. Le colmatage des venues d’eau dans le tunnel n’a reconstitué que partiellement le phénomène et une grande partie des eaux part en Suisse du côté de Vallorbe.
Il peut sembler surprenant que la nappe située sous le Mont d’Or alimente cette série de sources. Pour expliquer ce fait, Jean-Pierre Mettetal, hydrogéologue, évoque l’existence d’un vaste décrochement nord-sud qui partant de Vallorbe, intéresse les Hôpitaux-Neufs, la source du Bief Rouge, la Chapelle de Mijoux, la Combe où coulent la source puis le ruisseau de Fontaine Ronde, la Cluse-et-Mijoux, Pontarlier, Doubs et la N 57.
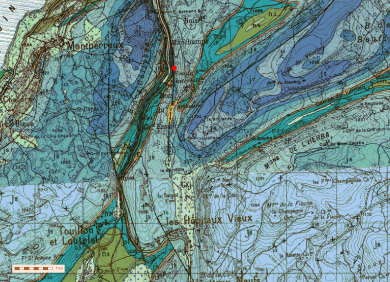
Le décrochement nord-sud passant par la source (cercle rouge)
En revanche, en ce qui concerne la grande source de Malbuisson, qui se situe dans le synclinal voisin, il n’existe pas à l’heure actuelle une explication à son rattachement à ce réseau souterrain.
Lors de la construction de la déviation de la RN 57 au-dessus des Hôpitaux-Vieux, les essais de traçage par coloration ont montré que les eaux se dirigeaient vers le nord et ressortaient au niveau d’un étang à proximité de la Jougnenat vers la pisciculture de Jougne. Ainsi les eaux provenant de l’est se dirigent vers le nord et celles provenant de l’ouest se dirigent vers le sud. Nous sommes ici sur la ligne de partage des eaux entre mer du Nord et Méditerranée.
Mais comment expliquer la périodicité du phénomène et son mécanisme ?
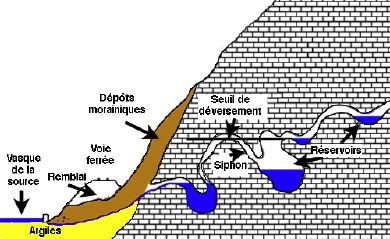
La présence de réservoirs karstiques successifs et superposés
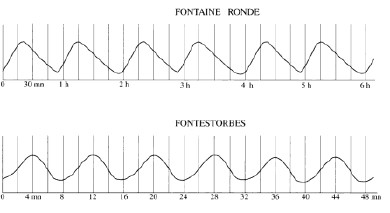
* Un réservoir amont (une salle souterraine creusée par l’eau dans la masse calcaire), alimenté par l’eau provenant du bassin versant ;
* Un conduit, dit de vidange, situé en position basse et emprunté par l’eau qui sort du réservoir pour rejoindre l’exutoire de la source ;
* Un conduit, dit de prise d’air, situé au-dessus et rempli d’air.

Fait capital pour le fonctionnement de la source, le hasard de l’érosion aurait placé les extrémités amont et aval des deux conduits côte à côte et sensiblement au même niveau. Ainsi, tant que le débit d’alimentation est suffisamment important, le niveau d’eau dans le réservoir amont maintient le conduit de prise d’air fermé. Dès lors, l’écoulement en charge dans le conduit de vidange se produit normalement au même rythme que le débit d’alimentation, exactement comme il le ferait si le conduit de prise d’air était absent, et la fontaine coule en continu.
En revanche, quand le débit d’alimentation baisse, à l’approche de l’étiage, il arrive un moment où il atteint 1,04 m³/s. À ce moment, l’extrémité amont du conduit de prise d’air, jusque là noyée et donc obstruée, se retrouve en partie hors d’eau, et aspire de l’air, ce qui produit une forte perte de charge dans le conduit de vidange et interrompt presque totalement l’écoulement : la première intermittence débute.
Le débit de vidange étant alors insignifiant par rapport au débit d’alimentation, le réservoir amont se remplit à nouveau. Mais pour réamorcer l’écoulement dans le conduit de vidange, il doit monter plus haut que le niveau où s’est déclenchée la première intermittence. C’est ce qui explique qu’une fois que l’écoulement se rétablit, le débit à l’exutoire de la source monte à 1,8 m³/s environ, soit un débit supérieur à 1,04 m³/s, débit d’alimentation à partir duquel apparaît le phénomène d’intermittence. Le débit de sortie étant notablement supérieur au débit d’entrée, le réservoir amont se vidange rapidement jusqu’à désamorçage de l’écoulement et le cycle recommence puis se succède à lui-même tant que le débit d’alimentation est insuffisant pour maintenir un écoulement constant, ce qui dure en général tout l’été et une partie de l’automne, avec simplement des interruptions occasionnelles de quelques jours à la suite d’orages qui font provisoirement remonter le débit d’alimentation.
Si, présenté de cette manière simplifiée, le mécanisme présente théoriquement une régularité parfaite, la réalité est un peu plus nuancée, en fonction de l’évolution du débit d’alimentation. Ainsi, au cours de la saison, le débit minimal varie entre 0,2 m³/s au début et 0,2 m³/s en fin d’été, quand le débit d’alimentation est au plus bas. De même, le débit maximal varie entre 1,80 et 1,68 m³/s, valeurs assez proches du module et qui font que la fontaine semble présenter un débit très abondant pour la saison si on s’y rend au bon moment du cycle.
La durée moyenne d’une intermittence, voisine d’une heure et partagée en environ 40 minutes de baisse et 20 minutes de hausse la plus grande partie de la saison, augmente sensiblement en fin d’étiage, quand les débits moyens sont au plus bas, car les réservoirs se remplissent moins vite.
Dans quelques cas assez rares au vu de l’hydrologie de la source, il peut arriver que les réservoirs ne puissent plus se remplir et que le débit d’alimentation puisse tout entier s’écouler de manière diphasique (eau + air) dans le conduit de vidange. Ceci se produit dès lors que le débit moyen journalier de la source descend vers 0,60 m³/s, soit 600 litres/s, ou en dessous, comme cela a été le cas lors de quelques étiages prolongés, en novembre 1973, octobre 2001 et janvier 2007. Cette valeur seuil avait été prévue presque exactement par les calculs effectués par A. Mangin qui avait trouvé 680 l/s.
En ce qui concerne Fontaine Ronde, une autre interprétation a prévalu dans le passé
Selon la légende, Amaury de Joux avait gardé des croisades une passion pour les longues chevauchées. Un jour en sortant du Château de Joux, la herse s’abattit brutalement coupant l’animal en deux sans que le cavalier s’en doutât. L’animal partit au grand galop. Mais la jument eut bientôt soif, s’arrêta à la source et but sans discontinuer jusqu’à l’assèchement du point d’eau. Amaury s’aperçut alors que l’eau qu’elle engloutissait sortait par la plaie sanglante. Épouvanté, il s’enfuit. Depuis l’animal est devenu invisible, mais il a toujours aussi soif. C’est lui qui revenant boire périodiquement, fait baisser le niveau de Fontaine Ronde.
Qui sait ? C’est peut-être la meilleure explication du fonctionnement de la source intermittente…
Remerciements à Jean-Pierre Mettetal et à Pierre Chauve pour leurs explications en ce qui concerne la source de Fontaine Ronde. La description du fonctionnement de la fontaine intermittente de Fontestorbes est due à Jean-Noël Salomon.
08:31 Publié dans Géologie-hydrogéologie-Climatologie | Tags : jura, karst, source intermittente, fontaine-ronde | Lien permanent | Commentaires (1) | Facebook | | |
Le Lémur couronné

Le Lémur couronné
Eulemur coronatus
par André Guyard
Le Lémur couronné, Eulemur coronatus, se trouve exclusivement dans le nord de Madagascar. Plus précisément, son aire de répartition s’étend du Cap d’Ambre au nord à la rivière Fanambana au sud, jusqu’à la ville d’Ambilobe et la rivière Sambirano à l’ouest et à la côte est. C'est à l'occasion d'une expédition organisée par le professeur Albignac, spécialiste des Lémuriens que nous avons eu l'occasion d'approcher de ce charmant animal.




Le Lémur couronné habite les forêts sèches du Cap d’Ambre et la Sakalava. Dans la forêt de l’Ankarana où nous l’avons observé, Eulemur coronatus se rencontre dans la canopée à la lisière des zones forestières dégradées. Habitué aux touristes, il n’hésite pas à s’approcher pour quémander de la nourriture.
C’est un animal de la taille d’un chat domestique. Il pèse environ 2 kg et mesure une trentaine de cm de long. La queue atteint 45-50 cm. Il tire son nom de la présence d’une couronne de poils de couleur orange sur le dessus de la tête.
Eulemur coronatus est polygame. Les accouplements se produisent en juin. La gestation dure environ 125 jours et les femelles mettent au monde un ou deux petits à la saison des pluies. La mère porte son bébé sous son ventre durant les trois premières semaines et l’allaite jusqu’à 5 à 6 mois. La maturité sexuelle est atteinte au bout de 20 mois. La longévité est de 27 ans en captivité.
Le Lémur couronné est diurne. Il est actif du lever au coucher du soleil avec une pause à midi qui peut durer quatre heures. Il a tendance à vivre en groupes de 5 à 6 individus. Les femelles sont dominantes. Comme chez tous les lémuriens, le grooming, c’est-à-dire caresses, toilette et épuçage mutuels, est important pour le développement et l’entretien des liens sociaux.
La communication entre individus implique éléments chimiques, sous forme de phéromones de marquage, éléments visuels : posture, expressions du visage, et échange de messages vocaux.
Le Lémur couronné est essentiellement frugivore, occasionnellement folivore, nectarivore, voire insectivore.
Ses prédateurs éventuels sont les rapaces et surtout le fossa, une espèce endémique de carnivore plantigrade à allure de félin, qui représente le plus grand prédateur terrestre des lémuriens de Madagascar.
Le Lémur couronné intervient dans son écosystème essentiellement comme un disséminateur de graines. Bien que protégé (liste rouge de l’UICN), il est souvent braconné pour sa viande. Mais la destruction de leur habitat constitue la principale menace pour les lémuriens couronnés. C’est pourquoi Eulemur coronatus a tendance à rester dans les limites de quatre réserves : la Forêt d’Ambre, le Parc national de la Montagne d’Ambre et la réserve de l’Ankarana.
La présence de Lémuriens à Madagascar constitue un important atout pour l’écotourisme malgache.
Photos et vidéo : André Guyard
Voir également sur ce même blog :
Le Lémur noir ou Lémur macaco,
Références
Inconnu. "Crowned Lemurs". Duke University Primate Center, http://primatecenter.duke.edu/animals/crowned/.
Harcourt, C., J. Thornback. 1990. Lemurs of Madagascar and the Comoros. Gland, Switzerland and Cambridge, UK : IUCN-The World Conservation Union.
Kappeler, P., J. Ganzhorn. 1993. Lemur Social Systems and Their Ecological Basis. New York : Plenum Press.
Mittermeier, R., W. Konstant, M. Nicoll, O. Langrand. 1992. Lemurs of Madagascar : An Action Plan for their Conservation 1993-1999. Gland, Switzerland : IUCN.
Nowak, R. 1999. Walker’s Mammals of the World, Sixth Edition. Baltimore and London : The Johns Hopkins University Press. 2008/05/25 04:07:11.507 GMT-4
Suter, M. 2000. "Eulemur coronatus", Animal Diversity Web. http://animaldiversity.ummz.umich.edu/site/accounts/information/Eulemur_coronatus.html
08:24 Publié dans Mammifères | Tags : lémur couronné, lémuriens, madagascar | Lien permanent | Commentaires (0) | Facebook | | |
05/10/2009
Le Courlis corlieu
 Le Courlis corlieu
Le Courlis corlieu
Numenius phaeopus
(Famille des Scolopacidés)
par André Guyard

Il fréquente les rives des cours d’eau et les prairies marécageuses



De son bec arqué, le Courlis sonde sédiments et graviers
22:02 Publié dans Limnologie-hydrobiologie, Ornithologie | Lien permanent | Commentaires (0) | Facebook | | |