



04/01/2017
Biodiversité et productivité de la forêt
Relations positives entre biodiversité
et productivité prédominantes
dans les forêts mondiales
La relation entre la biodiversité et la productivité des écosystèmes herbacés a été explorée en long et en large. En revanche, en ce qui concerne la flore arborescente, c'est-à-dire les forêts, les schémas sont beaucoup moins bien compris. Jusqu'à présent, il y avait eu peu d'expériences sur la diversité des arbres et le BPR forestier (effet de la biodiversité sur la productivité des écosystèmes) n'a été exploré que dans le cadre d'études d'observation à échelle régionale. Ainsi, la force et la variabilité spatiale de cette relation restaient inexplorées à l'échelle mondiale.
Pourtant, le BPR forestier se trouve à la base de la compréhension de la crise mondiale d'extinction et de ses impacts sur le fonctionnement des écosystèmes naturels. Le BPR forestier a été un sujet de recherche important dans l'écologie régionale au cours des dernières décennies, mais ce n'est que récemment qu'on a commencé à développer une perspective globale.

Parcelle monospécifique de charmes (forêt de Chaux, Jura)
Cliché DR
Dans le monde entier et sous toutes les latitudes, dans le cadre du projet international GFBI (Global Forest Biodiversity Initiative), un ensemble de 84 chercheurs (Liang et al. [2016]) ont amassé une somme impressionnante de données mondiales sur la forêt portant sur plus de 770 000 parcelles d'échantillonnage dans 44 pays.
Ils ont ainsi mis en évidence une relation positive et cohérente entre la diversité des arbres et la productivité des écosystèmes aux échelles du paysage, du pays et de l'écorégion. En moyenne, une perte de 10% de la biodiversité entraîne une perte de productivité de 3%. Cela signifie que la valeur économique du maintien de la biodiversité pour la productivité forestière mondiale est plus de cinq fois supérieure aux coûts de conservation globaux.

Forêt primaire tropicale (Bélouve, Réunion)
Cliché © André Guyard
Les forêts sont les plus importants dépôts mondiaux de la biodiversité terrestre, mais la déforestation, la dégradation des forêts, le changement climatique et d'autres facteurs menacent environ la moitié des espèces d'arbres dans le monde. Bien que des efforts considérables aient été déployés pour renforcer la préservation et l'utilisation durable de la diversité biologique des forêts dans le monde entier, les conséquences de cette perte de diversité constituent une grande incertitude pour les efforts internationaux de gestion et de conservation des forêts. La BPR de la forêt représente un chaînon manquant essentiel pour une évaluation précise de la biodiversité mondiale et une intégration réussie de la conservation biologique et du développement socioéconomique.

Parcelle de feuillus diversifiés au printemps (forêt de Thise)
Cliché © André Guyard

Parcelle de feuillus diversifiés en hiver (forêt de Chailluz)
Cliché © André Guyard
Les chercheurs du GFBI ont exploré l'effet de la richesse des essences forestières sur la productivité du volume d'arbres à l'échelle mondiale en utilisant des inventaires forestiers répétés de 777 126 parcelles d'échantillonnage permanentes dans 44 pays contenant plus de 30 millions d'arbres de 8737 espèces couvrant la plupart des biomes terrestres. Les résultats révèlent un effet positif cohérent de la biodiversité sur la productivité forestière à travers le monde, montrant qu'une perte continue de la biodiversité entraînerait un déclin accéléré de la productivité forestière mondiale.

Parcelle monospécifique de Douglas
Cliché © André Guyard
Le BPR montre une variation géospatiale considérable à travers le monde. Le même pourcentage de perte de biodiversité entraînerait un déclin relatif relatif (c'est-à-dire en pourcentage) de la productivité dans les forêts boréales de l'Amérique du Nord, de l'Europe du Nord-Est, de la Sibérie centrale, de l'Asie de l'Est et des régions éparses de l'Afrique centrale et méridionale. Dans l'Amazonie, l'Afrique de l'Ouest et du Sud-Est, le sud de la Chine, le Myanmar, le Népal et l'archipel malais, le même pourcentage de perte de biodiversité entraînerait un déclin absolu de la productivité.
En conclusion, la santé d'une forêt est liée à la diversité de ses arbres. Plus une parcelle forestière contient d'espèces, plus elle est productive — autrement dit, plus elle fabrique de matière végétale au mètre carré.
Mais de l'avoir démontré à pareille échelle devrait définitivement tordre le cou à l'idée que la parcelle la plus productive est celle où tous les arbres appartiennent à la même espèce. "De plus, la biodiversité est une assurance contre le changement climatique : si l'on a plus d'espèces, on a plus de chances que la forêt résiste bien au réchauffement."
Ces résultats mettent en évidence l'effet négatif de la perte de biodiversité sur la productivité forestière et les avantages potentiels de la transition des monocultures aux peuplements mixtes dans les pratiques forestières. La BPR découverte à travers les écosystèmes forestiers du monde entier correspond bien aux progrès théoriques récents, ainsi qu'aux études expérimentales et observationnelles sur les écosystèmes forestiers et non forestiers. Sur la base de cette relation, la perte constante d'espèces dans les écosystèmes forestiers dans le monde pourrait réduire considérablement la productivité forestière et donc le taux d'absorption du carbone forestier pour compromettre le puits mondial de carbone forestier.
La valeur économique de la biodiversité pour le maintien de la productivité forestière commerciale est estimée de 166 à 490 milliards de dollars par an. Bien qu'elle ne représente qu'un faible pourcentage de la valeur totale de la biodiversité, cette valeur est de deux à six fois supérieure à ce qu'elle coûterait pour mettre en œuvre efficacement la conservation à l'échelle mondiale. Ces résultats soulignent la nécessité de réévaluer l'évaluation de la biodiversité et les avantages potentiels de l'intégration et de la promotion de la conservation biologique dans la gestion des ressources forestières et les pratiques forestières dans le monde entier.
Source :
Liang et al. (2016).— Positive Biodiversity–Productivity Relationship Predominant in Global Forests Science 14 oct. 2016 : Vol. 354, n° 6309, DOI: 10.1126 / science.aaf8957
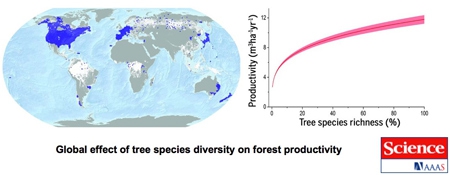
Effet global de la diversité des
essences forestières sur la productivité forestière
Les données obtenues au sol provenant de 777 126 parcelles d'échantillonnage permanentes de la diversité biologique des forêts (zone bleu foncé), qui couvrent une partie substantielle de l'étendue forestière mondiale (en blanc), révèlent des relations positives. La ligne rouge concave avec des bandes roses représentant un intervalle de confiance de 95%).
La filière bois en France
(Vidéo enquête de Samuel Ruffier et Paul Aurélien Combre de France 3 Bourgogne—Franche-Comté)
11:34 Publié dans Botanique, Environnement-Écologie | Tags : biodiversité et productivité de la forêt, global forest biodiversity initiative | Lien permanent | Commentaires (0) |  Facebook | | |
Facebook | | |
05/11/2016
Automne 2016
Zoom sur l'automne
par Dominique Delfino
Photographe naturaliste
C'est avec un regard un peu particulier que je partage ce cliché de fin de saison.
Juste pour rappeler que l'automne est passé comme une lettre à la poste…
Cette image aux perspectives fuyantes est le résultat ''d'un coup de zoom'' réalisé lors de la prise de vue dans ce sous-bois.
Une façon peut-être de faire étinceler une dernière fois les lumières d'automne avant qu'elles ne laissent la place aux rayonnements spécifiques des jours d'hiver.

15:00 Publié dans Botanique, Environnement-Écologie, Photographes naturalistes et scientifiques | Tags : dominique delfino, photographe naturaliste et animalier, automne | Lien permanent | Commentaires (0) | Facebook | | |
04/11/2016
Potirons arbustifs à Brognart ?
Potirons arbustifs à Brognart ?
par Dominique Delfino
Photographe naturaliste et animalier
L'automne dévoile quelquefois des surprises étonnantes et quelle ne fut pas celle de mon voisin Daniel Brandt lorsqu'il découvrit quelques potirons accrochés aux branches d'un arbre en bordure de sa propriété.
Bien dissimulés dans la végétation au cours de l'année, ils sont passés totalement inaperçus jusqu'à ce que la chute des feuilles laisse apparaître une imposante masse dominant par ailleurs le bord de la route.
Le compost au coin du jardin a tout simplement permis aux graines de l'année précédente de germer et de se développer en rampant le long du tronc d'arbre pour s'épanouir en altitude.
Une ''cueillette'' qui nécessitera des moyens hors du commun pour récolter ces légumes d'automne avec l'aide de Daniel Nusbaumer, agriculteur en retraite, qui dut mettre en œuvre un tracteur et une grande échelle.
Bilan : un potiron d'environ sept kilos qu'il valait mieux ne pas prendre sur le pare-brise ou sur la tête lors d'une petite balade champêtre !

Cliché © Dominique Delfino
10:12 Publié dans Botanique, Photographes naturalistes et scientifiques | Tags : cucurbitacées, potion, brogniart, pays de montbéliard, dominique delfino, photographe naturaliste et animalier | Lien permanent | Commentaires (0) | Facebook | | |
19/09/2016
Le polypore soufré
Sous la lumière du Polypore
par Dominique Delfino
Photographe naturaliste et animalier
Un polypore soufré s'accroche au tronc de ce vieux cerisier bordant le canal de Haute-Saône à Brognard. La lumière réfléchie par ce champignon particulièrement bien exposé est flamboyante.
Le Polypore soufré est un parasite des blessures de nombreux arbres. Il se rencontre du printemps à l'automne, sur l'écorce de troncs verticaux ou couchés de nombreux feuillus et arbres fruitiers (rarement sur les conifères).
Très jeune, ce champignon est considéré comme comestible et apprécié dans certains pays comme les États-Unis.
Mais le régal est avant tout pour les yeux à chaque fois que j'emprunte le chemin où s'expose le Polypore.
Les rayons lumineux qui le traversent à contre-jour donnent à sa couleur un éclat renforcé par son développement à hauteur d'homme et que mon objectif grand angle permettra de saisir sous cette perspective.

Cliché © Dominique Delfino
06:47 Publié dans Botanique, Mycologie, Photographes naturalistes et scientifiques | Tags : dominique delfino, photographe naturaliste et animalier, champignons, polypore soufré | Lien permanent | Commentaires (0) | Facebook | | |
17/09/2016
Liste hiérarchisée des espèces végétales invasives de Franche-Comté : rapport complet
Liste hiérarchisée des espèces végétales invasives de Franche-Comté
Le Conservatoire botanique national de Franche-Comté publie un rapport complet sur les espèces végétales invasives de Franche-Comté.
Dix ans après la première liste (2006), la méthodologie de classement des espèces et la terminologie employée pour la catégorisation ont été revues, dans un souci notamment de recherche de cohérence avec les autres listes établies sur le territoire national par chacun des conservatoires botaniques nationaux. En outre, le classement des espèces s’est essentiellement appuyé cette fois sur la consultation de la base de données Taxa, en raison de la progression significative du nombre et de la qualité des observations d’espèces exotiques depuis dix ans. Les actions pressenties pour chacune des catégories d’espèces exotiques identifiées sont proposées.
13:59 Publié dans Botanique, Environnement-Écologie, Patrimoine franc-comtois et jurassien | Tags : espèces invasives, franche-comté, conservatoire botanique de franche-comté | Lien permanent | Commentaires (0) | Facebook | | |
29/07/2016
Dégâts forestiers : le cerf blanchi

08:20 Publié dans Actualité des Sciences, Botanique, Environnement-Écologie, Mammifères | Tags : cerf, dégâts forestiers, est républicain | Lien permanent | Commentaires (0) | Facebook | | |
14/07/2016
Le chêne de Belverne
Le chêne de Belverne
par Dominique Delfino
Photographe naturaliste et animalier
Ce chêne est tout simplement majestueux, superbe et ma fascination pour les vieux arbres est à son comble lorsque je découvre en lisière de forêt ce sujet signalé par l'un de mes amis sur son terrain à Belverne (70).

Hormis les centaines années que compte probablement son âge, la particularité de ce chêne réside dans le développement des deux troncs que comporte l'arbre. Comme pour mieux consolider cette architecture végétale, une branche transversale soudée naturellement par le temps, relie les troncs comme pour mieux renforcer l’équilibre de ce monument naturel.

Comment cette branche au cours de sa lente croissance a t-elle pu finalement au contact du tronc voisin se greffer et finir par se souder pour réaliser ce renfort naturel ?
Une particularité que l'on peut attribuer au hasard mais qui en tout cas confirme bien l'expression ''le hasard fait bien les choses''.
10:23 Publié dans Botanique, Photographes naturalistes et scientifiques | Tags : dominique delfino, photographe naturaliste et animalier, chêne, belverne 70 | Lien permanent | Commentaires (0) | Facebook | | |
14/06/2016
Coquelicots
Ils flottent au gré du vent
par Dominique Delfino
Photographe naturaliste
Qui n'a pas remarqué ce superbe champ de coquelicots en contrebas de la route face à l'hypermarché entre Montbéliard et Bethoncourt ?
Ce terrain demeuré en friche, retrouve une dynamique végétale naturelle, offrant à la flore la possibilité de s'épanouir librement.
Petits mammifères, insectes et oiseaux, dont certaines espèces sont protégées, trouvent ici le refuge pour s'y installer le temps d'une saison liée à la reproduction.
C'est la couleur rouge pourpre qui s'est imposée durant plusieurs semaines, transformant cet espace en campagne colorée au cœur d'un milieu urbain dense.
Malgré la météo capricieuse de ce printemps qui se fait attendre, le coquelicot nous fait songer à l'été, au soleil et nombreux sont les admirateurs qui se sont laissé tenter par quelques photographies.
Les fleurs dansantes et souples s'agitent au moindre souffle du vent, les pétales fripés accentuent la fragilité du coquelicot, un spectacle dont il faudra profiter avant que ne s'éteignent ces étincelles de lumière dans le paysage.

Cliché © Dominique Delfino
09:56 Publié dans Botanique, Environnement-Écologie, Photographes naturalistes et scientifiques | Tags : dominique delfino, photographe naturaliste et animalier, coquelicots | Lien permanent | Commentaires (0) | Facebook | | |
02/05/2016
Mosaïque de colza
Mosaïque de colza
par Dominique Delfino
Photographe naturaliiste et animalier
En ce printemps 2016, qui n'a pas remarqué la métamorphose de couleurs que subissent nos campagnes depuis quelques semaines ?
Le colza semble avoir pris le dessus au sein des parcelles agricoles et sa floraison se maintient particulièrement bien dans le temps cette saison. En revanche, le maïs n'est pas sorti du sol car le début d'année trop pluvieux ne permettait pas le travail de la terre et qu'il n'a pas encore été semé m'explique un agriculteur.
Je profite d'une très belle lumière de début de soirée en ce dimanche 8 mai pour prendre de la hauteur et redécouvrir à vol d'oiseau ces paysages dans les environs de Montbéliard.
Très vite, les parcelles d'un jaune lumineux se dessinent à l'horizon de la campagne environnante mais s'affichent également aux portes de l'agglomération, contrastant entre agriculture et urbanisme.
C'est l'immensité de certaines parcelles qui surprend par leur taille tel un gigantesque damier vu du ciel. L'exploitation de terrains à une échelle bien moindre offre des cultures qui s'imbriquent les unes dans les autres comme en témoigne cette image de mosaïque végétale.

Cliché © Dominique Delfino
22:26 Publié dans Botanique, Environnement-Écologie, Photographes naturalistes et scientifiques | Tags : dominique delfino, photographe naturaliste et animalier, colza, paysage printanier | Lien permanent | Commentaires (0) | Facebook | | |
29/11/2015
Sorties naturalistes en Franche-Comté
Sorties naturalistes en Franche-Comté
Balades nature de l'Office du Tourisme
et des Congrès de Besançon
Propositions de balades nature du premier semestre 2016
Sur inscription préalable et indispensable, auprès de l'Office du Tourisme et des Congrès de Besançon (03 81 80 92 55) ; annulations possibles ou sorties différées si trop peu d'inscrits ou en fonction des aléas de la météorologie.
DÉCOUVREZ LES SENTIERS AMÉNAGÉS SUR LE GRAND BESANÇON
18 boucles pédestres, 115 km de circuits (hors liaisons) :
8 boucles de difficulté familiale, 8 boucles de difficulté moyenne, 2 boucles de difficulté sportive, 6 boucles VTT
100 km de circuits (hors liaisons) : 2 circuits de difficulté moyenne (bleu), 3 circuits de difficulté difficile (rouge), 1 circuit de difficulté très difficle (noir)
Pour découvrir les circuits pédestres et VTT, imprimez les fiches, télécharger les fichiers GPS, cliquez ICI
04:03 Publié dans Botanique, Environnement-Écologie, Mammifères, Patrimoine franc-comtois et jurassien | Lien permanent | Commentaires (0) | Facebook | | |
22/11/2015
Curieux de nature dans la Reculée des Planches
Curieux de nature dans la Reculée des Planches

06:18 Publié dans Botanique, Environnement-Écologie, Livres, Ornithologie, Patrimoine franc-comtois et jurassien | Lien permanent | Commentaires (0) | Facebook | | |
06/10/2015
Permaculture
La permaculture
La permaculture est une méthode systémique et holistique de conception d'habitats humains et de systèmes agricoles inspirée de l'écologie naturelle (biomimétisme) et de la tradition. Elle n'est pas un mode de pensée mais un mode d'agir qui prend en considération la bio-diversité des écosystèmes1,2. En outre, elle vise à créer une production agricole durable, très économe en énergie (travail manuel et mécanique, carburant...) et respectueuse des êtres vivants et de leurs relations réciproques, tout en laissant à la nature « sauvage » le plus de place possible1,2.
Cette conférence a pour but de donner une idée du stage PDC de permaculture qui aura lieu du 6 au 18 juin 2016 au Lycée agricole de Valdoie.

08:28 Publié dans Actualité des Sciences, Botanique, Nature & philosophie, Nature et santé, Pollution | Tags : permaculture, bernard alonso, lycée agricole de valdoie | Lien permanent | Commentaires (0) | Facebook | | |
08/09/2015
Vergers vivants recrute un Directeur (secteur Montbéliard - 25)
Vergers vivants
association membre de FNE Franche-Comté
recrute un Directeur
(secteur Montbéliard - 25)
 SECRETARIAT
SECRETARIAT
MEFC - 7, rue Voirin - 25000 Besançon
Tél. 03 81 80 92 98 - http://fne-franche-comte.fr/
Association agréée au titre de la loi du 10 juillet 1976 relative à la Protection de la Nature
---------- Message transféré ----------
De : Nicolas LAVANCHY - Vergers Vivants <n.lavanchy@vergers-vivants.fr>
Date : 8 septembre 2015 18:38
Objet : offre de poste DIRECTEUR/TRICE
À :
Bonjour,
Après 5 ans d'implication au sein de Vergers Vivants, je vais quitter mes fonctions de directeur en cette fin d'année.
Ainsi, l'association recrute une personne pour prendre le relais à compter de janvier 2016. Vous trouverez en pièce jointe une offre d'emploi à diffuser pour le poste de DIRECTEUR / DIRECTRICE.
Merci d'avance et au plaisir de vous croiser bientôt !
Cordialement,
--
 Nicolas LAVANCHY
Nicolas LAVANCHY
VERGERS VIVANTS
La Damassine
23, rue des Aiges
25230 VANDONCOURT
Tél. 03 81 37 82 26
www.vergers-vivants.fr


09:20 Publié dans Actualité des Sciences, Botanique, Nature et santé, Patrimoine franc-comtois et jurassien | Tags : vergers vivants, fne-franche-comte, la damassine | Lien permanent | Commentaires (0) | Facebook | | |
03/03/2015
Le séquoia président du parc des séquoias de Californie
Le séquoia président du parc des séquoias de Californie
L'âge du "Président" du parc des séquoias de Californie, ce séquoia géant qui s'élève à 247 pieds de haut (80 m) est estimé à plus de 3.200 ans soit 1200 ans avant J.-C.

© National Geographic
Le tronc du Président mesure 27 pieds de diamètre et son feuillage se pare de 2 milliards d’aiguilles.

© National Geographic
En raison de sa taille incroyable, cet arbre n'avait jamais été photographié dans son intégralité.

© National Geographic
Une équipe de photographes du National Geographic a travaillé avec des scientifiques pour essayer de créer la première photo qui montre le Président dans toute sa splendeur.

© National Geographic
Ils ont dû grimper à l'arbre avec des poulies et leviers, et ils ont pris des milliers de photos.

© National Geographic
Parmi celles-ci, ils en ont choisi 126 et les ont assemblées pour obtenir cette incroyable photo du Président.

© National Geographic
09:09 Publié dans Botanique, Environnement-Écologie, Nature & philosophie | Lien permanent | Commentaires (0) | Facebook | | |
24/02/2015
SHNPM : calendrier 2015




12:02 Publié dans Actualité des Sciences, Botanique, Environnement-Écologie, Mycologie | Lien permanent | Commentaires (0) | Facebook | | |
12/02/2015
Société botanique de Franche-Comté : Sorties et animations année 2015
Société botanique de Franche-Comté : Sorties et animations année 2015


14:08 Publié dans Botanique | Tags : franche-comté, botanique | Lien permanent | Commentaires (0) | Facebook | | |
09/12/2014
La chalarose du Frêne
 La chalarose du Frêne
La chalarose du Frêne
(dernière mise à jour : 30 /03/2019)
Un champignon menace nos frênes, rappelant les ravages subis par l'orme avec la graphiose au siècle dernier. Or le frêne représente une essence d'une grande importance écologique et économique qui est ainsi menacée dans son existence.
Voir ici une vidéo de France 3 Bourgogne-Franche-Comté
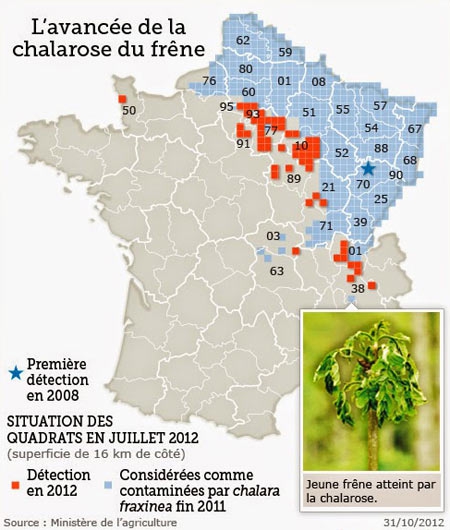
À la fin du siècle dernier, une nouvelle maladie, la chalarose est apparue sur le frêne dans l'est de l'Europe. En fait, elle serait originaire d'Asie via la Pologne ou les pays baltes. Cette maladie provoque des symptômes variés à différents niveaux de l’arbre. Sur les jeunes sujets, les premiers symptômes s’observent facilement. Ils se manifestent par une pousse tardive des feuilles et une chute prématurée (dès mi-août), avec apparition de branches sèches au niveau de la cime. Les jeunes rameaux se flétrissent puis se nécrosent. Des chancres se forment et l'on assiste à une descente de la cime. Chez les jeunes sujets, l'atteinte est le plus souvent mortelle. Au niveau du collet, la maladie peut initier des nécroses, qui peuvent être envahies par l’armillaire, ce champignon opportuniste qui se développe à l'automne sur les souches. Chez les sujets adultes, la dégradation semble lente et le taux de mortalité reste, pour l’instant, faible. Cependant leur aspect est nettement dégradé. Leur fonction d’ombrage ou d’ornement s’en trouve compromise.

Le frêne de droite est frappé par la chalarose
(cliché DR)
En 2006 une équipe de scientifiques polonais a pu l'isoler, identifier la pathologie due au champignon Chalara fraxinea d'où le nom donné à la maladie. Selon Leslie Constantin, technicienne de l’association des communes forestières du Jura, Chalara fraxinea “provoque notamment une nécrose de l’écorce, qui constitue une protection de l’arbre contre les parasites, le rendant donc plus vulnérable”. Une nécrose qui peut atteindre le pied de l’arbre, provoquant la pourriture du bois et l’exposition à d’autres parasites pouvant accélérer la mortalité des arbres.

Les jeunes frênes atteints subissent d'abord un flétrissement
Rapidement les pays voisins ont détecté également la maladie repérée jusqu'en Finlande, en Allemagne, Autriche, Suisse, et depuis 2008, en France, où elle semble bien établie.
L'agent de la chalarose, Chalara fraxinea est un hyphomycète, champignon ascomycète microscopique imparfait au stade asexué (anamorphe) indétectable à l'œil nu. Depuis peu, on connaît son stade parfait (téléomorphe) autrement dit le champignon au stade sexué : Hymenoscyphus pseudoalbidus (Baral. Queioz et Hosoya, 2014).
Au stade sexué, ce champignon peut être repéré assez facilement. Au printemps, il suffit de trouver une frênaie contaminée et de repérer au sol des pétioles des feuilles de l'an passé. Lorsque ces pétioles tombés sont totalement noircis, on peut observer de petits champignons blancs qui ressemblent chacun à un petit bouton monté sur un petit pied (jusqu'à 5 mm de diamètre pour un peu moins en hauteur : une loupe est quand même souhaitable pour mieux observer ce petit hyphomycète).

Ces petits carpophores vont émettre des spores qui seront disséminées par le vent. Le vent est en effet le vecteur principal de contamination. La densité de spores est encore importante à 500 mètres de la source, d’où des contaminations possibles des peuplements de proche en proche à l’échelle d’une collectivité. Le transport du bois peut être une source de contamination ainsi que la plantation de sujets provenant d’une zone contaminée.
Attention toutefois à bien identifier la maladie, car la mortalité des rameaux n'est pas spécifique à la chalarose et des facteurs climatiques extrêmes peuvent aussi affaiblir l'arbre. D'ailleurs il n'est pas impossible que ces facteurs rentrent en jeu dans l'action de la maladie. Pour être plus sûr de l'identification, il conviendra de repérer des nécroses corticales présentes à la base des rameaux morts, fréquemment sur les petits rameaux latéraux.
On a longtemps cru ce champignon cantonné dans les départements d'Alsace-Lorraine, Franche-Comté, en progression lente vers l'ouest ou le sud. Ce sont d’abord la Haute-Saône puis le Pas-de-Calais qui ont été touchés, mais la chalarose est également implantée depuis longtemps aussi en Champagne-Ardenne et Bourgogne et aussi présente dans l'ouest et le sud de la France. Disons que sa carte de répartition est provisoire et en cours d'élaboration (voir ci-dessous la répartition en 2016).
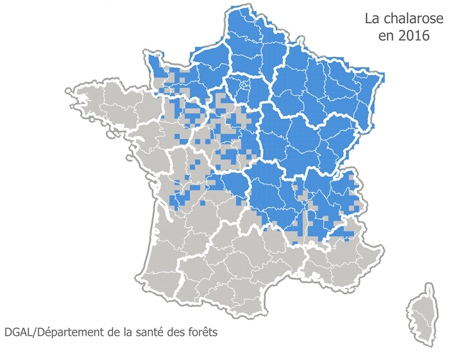
Dans le Jura, où le frêne représente 4 % du volume des forêts, le parasite est apparu, il y a moins de cinq ans : les premiers signes ont été identifiés en 2010. Il est aujourd’hui présent partout dans le département où le frêne pourrait être totalement décimé, comme ce fut le cas pour l’orme confronté à la graphiose dans les années 80.
Dans le Doubs, il est identifié dans différentes localités. Par exemple, le bouquet de frênes qui ombrage l'aire de jeux de la commune de Thise, devra être abattu et brûlé, car, de toute évidence, il est frappé par la chalarose.
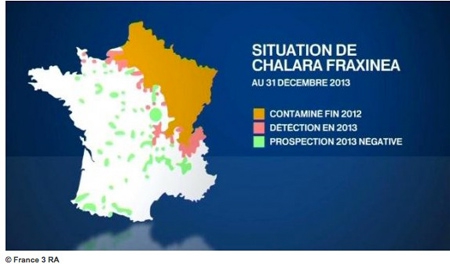
En 2002, le ministère de l’Agriculture se voulait encore optimiste : “ Les mortalités arrivent tardivement, elles restent pour l’instant faibles, la dégradation des frênes paraît lente et certains semblent aller mieux ”, indiquait en conclusion un rapport du département de la santé des forêts sur l’avancée de la chalarose du frêne en France. “S’il est indiscutable que la maladie progresse sur le territoire français comme européen, les facteurs d’occurrence et les conséquences de la maladie sont aujourd’hui encore mal connus. Les premiers constats alarmistes que renvoyaient les symptômes préoccupants dans les houppiers et la rapidité de propagation de la maladie sur le territoire laissaient entrevoir un avenir très incertain pour le frêne. Les résultats des différents suivis installés, en France par le DSF, mais également dans d’autres pays d’Europe, atténuent cette prévision.
Les mortalités arrivent tardivement, elles restent pour l’instant faibles, la dégradation des frênes semble lente, certains arbres ne montrent pas ou peu de symptômes, d’autres se stabilisent ou semblent aller mieux... Cependant, si les arbres adultes peuvent endurer la maladie, cette dernière est souvent létale pour les jeunes frênes et compromet la régénération des peuplements. En France, les dégâts les plus significatifs apparaissent liés aux nécroses au collet. Ces dernières semblent directement liées aux mortalités et aux arbres très dépérissants. Les pays d’Europe contaminés s’accordent sur le fait que si aucun arbre ne semble totalement résistant à la maladie, certains individus reste en relativement bon état sanitaire malgré une forte pression d’inoculum. Cela suggère que ces arbres pourraient être à l’origine d’une population tolérante à la chalarose.”
Or, depuis, la situation s’est grandement détériorée et là où le même rapport laissait entrevoir l’existence d’une population "tolérante" à la maladie, les constatations sur le terrain amènent à un tout autre diagnostic : “ N’importe qui peut voir que les frênes dépérissent ” assure Michel Bourgeois, le président de l’association des communes forestières du Jura.
En 2014, Serge Outrey, le conseiller général de Nozeroy, a alerté les services du département du Jura de l’avancée de la chalarose dans son canton : “L’an dernier, on a vu les premiers signes sur le Premier plateau. La maladie est chez nous depuis ce printemps”. Dans cette partie du département plus qu’ailleurs, on trouve surtout le frêne dans les haies ou en bord de route. Et c’est à cet endroit que les arbres malades peuvent devenir dangereux : “J’estime à une centaine le nombre de frênes à abattre dans le canton pour éviter les chutes de branches sur la chaussée” indique l’élu.
Selon Bruno Guespin, technicien à l’ONF et correspondant du pôle santé des forêts, “la chalarose est présente sur l’ensemble du Jura”. Mais il semble qu’on ait décidé de ne pas trop affoler les collectivités de manière à éviter les coupes massives et une chute des cours.
Certes, le frêne n’est pas le bois le plus répandu dans les forêts jurassiennes et sa disparition aurait surtout des conséquences paysagères. À ce stade, on estime que l’impact économique immédiat de la chalarose sera limité : le frêne ne représente que 4 % des volumes de bois commercialisés chaque année par les communes forestières.
Mais le phénomène devrait marquer la campagne d’abattage de 2015 : confrontées à l’avancée de la maladie, les communes pourraient être contraintes de se lancer dans un abattage massif. Cela provoquerait une arrivée importante de bois de chauffage sur le marché, ce qui fait craindre une baisse du cours des autres bois de chauffage, le hêtre et le chêne, dont les communes tirent l’essentiel de leurs revenus forestiers. Pour l’heure, on commence seulement à mesurer l’ampleur des dégâts, sans être capable d’en chiffrer les conséquences économiques. “Aucun recensement précis n’a encore été entrepris par les communes” convient le président des communes forestières". Mais Bruno Guespin a déjà une vision très claire de ce qui devrait arriver : “La seule question aujourd’hui est de savoir s’il y aura des survivants. Si les arbres qui sont sains aujourd’hui le seront dans cinq ans.” Car chez les scientifiques, on en est, en effet, à espérer qu’il existe une variété de frênes qui soit capable de résister à la chalarose…

Si les arbres adultes peuvent endurer la maladie, celle-ci est souvent fatale pour les jeunes frênes et compromet la régénération des peuplements.
En fait, les arbres desséchés représentent un dangers pour les promeneurs. L’Office national des forêts vient d’annoncer la fermeture au public de cinq forêts domaniales des Hauts-de-France. L'ONF craint les accidents. C’est pour cette raison que quatre massifs du Pas-de-Calais (Boulogne, Desvres, Hardelot et Vimy) et celui de Nieppe dans le Nord vont être fermés au public dès le 15 septembre 2016 et au moins jusqu’à l’été 2017. Les forestiers constatent en effet un affaiblissement des frênes tel que les chutes d’arbres et de branches sont à craindre le long des 350 kilomètres de chemins de randonnées parcourant ces forêts. Il va falloir près d’une année pour que les fonctionnaires de l’ONF inspectent systématiquement les arbres au voisinage des chemins et abattent les plus dangereux.
Seul point positif, la qualité des grumes ne semble pas altérée par le champignon. À ce jour, aucun moyen de lutte efficace n’est encore connu et les peuplements de frênes dans l’ensemble de l’Europe sont fortement menacés. La situation est donc préoccupante. Les plus anciens se souviennent assurément des ravages provoqués sur l'orme au siècle dernier par une autre maladie : la graphiose, un champignon (Ophiostoma ulmi) transmis par le scolyte de l'orme (Scolytus scolytus). Les dégâts ont été considérables et l'on a cru que les ormes seraient littéralement rayés de la carte. Même si on peut encore observer cette essence dans nos contrées, notons que les arbres ne deviennent pas si gros qu'autrefois. On ne peut pas s'empêcher de faire le parallèle avec le frêne, même si le recul nous manque un peu.

Au bout de quelques mois, le flétrissement est total
Cliché DR
Où est la solution ?
Pour l'instant, on se contente de brûler les sujets malades pour limiter la prolifération.
- La solution chimique ?
Un nombre considérable de maladies pathogènes, parmi lesquels les champignons, sont présents sur de multiples essences et font partie du cycle de la vie de celles-ci. L'Homme a souvent eu recours à des méthodes draconiennes comme les pesticides pour éradiquer les pathogènes des végétaux. L'agriculture intensive est un sinistre exemple où l'Homme finit par empoisonner l'ensemble des chaînes alimentaires. La solution, si elle existe, est probablement naturelle.
C'est pourtant fondée sur la chimie la voie choisie par des chercheurs britanniques. Les spécialistes de l’environnement Natural Ecology Mitigation travaillent sur l’élaboration d'une solution à base de fongicides avec la cellule de recherche sylvicole de la Forestry Commission, le International Pesticide Application Research Consortium et les chercheurs du Département des Sciences de la vie du Imperial College.
Tim Mott, directeur de Natural Ecology Mitigation, explique le déroulement du projet : "Le produit a été formulé et breveté ; les premiers essais au laboratoire ont été concluants. Nous menons la contre-attaque scientifique engagée contre la maladie des arbres et pensons que l’heure est venue d’accélérer la recherche en laboratoire et de multiplier les essais sur le terrain, afin d’être prêt à faire face à la nouvelle menace."
Le produit proposé par Natural Ecology Mitigation a été baptisé CuPC33 – solution biocide / fongicide à base de sulfate de cuivre et d’autres minéraux. L’utilisation du cuivre dans le traitement des maladies fongiques qui sévissent dans les foyers et jardins n’est pas récente ; plusieurs produits à base de cuivre ont déjà été commercialisés avec l’approbation des autorités sanitaires du Royaume-Uni.
Les essais en laboratoire montrent la grande capacité du produit à maîtriser le champignon à l’origine des maladies des arbres. Des essais en serres réalisés sur Silwood Park Campus ont montré l’innocuité pour les arbres du produit pulvérisé ou injecté.
Selon les chercheurs, le CuPC33 pourrait être disséminé dans les forêts touchées par pulvérisation ou sous forme de brume dense médicamenteuse qui se déposerait sur les feuilles et les branches. Grâce à une technologie d’atomisation du liquide en gouttelettes très fines, ils pensent que 10 litres de CupPC33 dilué suffiraient pour traiter un hectare de forêt, à raison de moins de 60 pence par litre (100 pence = une livre sterling). La main-d’œuvre et les machines seraient à l’origine de la majeure partie du coût total du traitement.
Simon Leather, professeur invité du Département des Sciences de la vie du Imperial College London, supervise la recherche sur le terrain à travers Imperial Consultants : "Nous espérons pouvoir mettre au point plusieurs moyens d’appliquer le CuPC33, en fonction des différents types de champignons, de maladies bactériennes et d’espèces d’arbres. Par exemple, une formule de CuPC33 à pulvérisation manuelle permettrait aux jardiniers de traiter leurs plantes décoratives et pourrait contribuer aux efforts visant à limiter la propagation de la maladie."
Le CuPC33 est capable de détruire plusieurs des champignons pathogènes qui menacent les arbres et les plantes, dont Pseudomonas syringae (plus connue sous le nom de chancre suintant du marronnier d’Inde) et Phytophthora ramorum, à l’origine de la mort subite du chêne qui touche le chêne et le mélèze. L’équipe de Natural Ecology Mitigation pense que le CuPC33 sera tout aussi efficace dans le traitement du dessèchement des pousses du frêne.
"Les arbres adultes ne peuvent pas être remplacés pendant plusieurs générations, mais traités à temps, les arbres malades peuvent encore être sauvés pour nos enfants et leurs familles. Les efforts mis en œuvre pour endiguer la propagation de la maladie en favorisant la santé des arbres par le recours à des éléments fertilisants, ont donné peu de résultats positifs et semblent voués à l’échec. Des mesures rapides doivent être prises dès maintenant, pour éliminer toutes les maladies fongiques envahissantes" conclut Tim Mott.

Cliché DR
- La lutte biologique par virus fongiques : un procédé plus élégant
Un projet de coopération entre la Suisse et la Lituanie peut engendrer de nouvelles possibilités de lutte contre cette maladie. Dans ce projet, les populations épidémiques (suisses) et post-épidémiques (lituaniennes) sont étudiées quant à leur virulence, leur diversité génétique et la présence de virus fongiques. Il s'agit aussi de déterminer dans quelle mesure les virus fongiques identifiés peuvent être utilisés pour une lutte biologique contre la maladie.
Quels sont les points forts du projet ?
- Comparaison de la diversité génétique et de la virulence entre les populations post-endémiques et endémiques de Chalara fraxinea.
- Recherche des virus fongiques de Chalara fraxinea par des analyses métagénomiques.
- Caractérisation des virus fongiques et détermination de leur incidence sur les différentes populations de l'agent pathogène.
- Étude de l'effet des virus fongiques identifiés sur leur hôte Chalara fraxinea et évaluation de leur utilité potentielle dans la lutte biologique contre la maladie
- Publication des résultats de recherche à l'intention de la science et d'un large public.
Quels sont les résultats attendus ?
- Données sur l'épidémiologie et la génétique de population de Chalara fraxinea l'agent pathogène de la chalarose du frêne.
- Clarification de l'hypothèse selon laquelle la virulence d'agents pathogènes invasifs diminue avec le temps.
- Renseignements sur la présence possible de virus fongiques encore inconnus dans Chalara fraxinea.
- Connaissances sur l'importance biologique et écologique des virus fongiques trouvés.
- Évaluation de l'efficacité potentielle de ces virus dans le cadre de la lutte biologique contre la maladie.
Ce projet est financé dans le cadre du programme de coopération helvético-lituanien visant à réduire les disparités économiques et sociales au sein de l'Union européenne élargie : projet n° CH-3-ŠMM-OV12.
Partenaires du projet :
— Nature Research Centre, Laboratory of Phytopathogenic Microorganisms, Institute of Botany, Vilnius, Lituanie.
— Institut fédéral de recherches sur la forêt, la neige et le paysage, Birmensdorf, Suisse.
Durée du projet : novembre 2012 à avril 2016.
Une autre parade possible : la sélection d’individus qui ne sont pas affectés par le champignon.
Car, heureusement, les frênes sont plus ou moins résistants à la maladie. Identifier cette résistance génétique, c’est le premier objectif du programme « Chalfrax » mené par le Centre national de la propriété forestière (CNPF). Il s’agit de constituer une population de frênes tolérant la maladie en préservant la biodiversité génétique de l’espèce.
Le second objectif est de rechercher de nouveaux modes de gestion sylvicole afin de contenir la maladie. Enfin, les forestiers cherchent des alternatives à l’exploitation du frêne. Il n’est en effet plus possible de s’en tenir à la repousse dynamique de cette espèce pionnière qui reconquiert naturellement — comme le chêne — les terres qui lui sont favorables. Si les arbres adultes — d’un diamètre supérieur à 45 centimètres — peuvent encore résister à la maladie, ce n’est pas le cas des jeunes individus qui sont très fortement affectés. Le CNPF estime ainsi que les jeunes peuplements de frênes ne sont plus rentables. On envisage de remplacer cette espèce par des aulnes, noyers, chênes, érables…
Consulter également :
http://www.fredon-bourgogne.com/
FREDON FRANCHE-COMTE École-Valentin N° 03 81 47 79 20
Liens :
http://www.wsl.ch/fe/biodiversitaet/projekte/eschentriebs...
13:58 Publié dans Botanique, Environnement-Écologie, Mycologie, Nature et santé, Parasitologie | Tags : chalarose du frêne, champignons, hyphomycètes, ascomycètes, phytopathologie, sylviculture, forêts | Lien permanent | Commentaires (2) | Facebook | | |
23/09/2014
La forêt comtoise dans l'histoire
 Il était une fois... la forêt comtoise
Il était une fois... la forêt comtoise
Les chercheurs francs-comtois du laboratoire Chrono-environnement de l'Université de Franche-Comté s'intéressent à l'histoire de la forêt comtoise. Ces quelques lignes reproduisent in extenso un article paru dans "En Direct" n° 254 septembre-octobre 2014.
Hantée par les ogres et les loups dans les légendes, la forêt était en réalité habitée de bûcherons, de charbonniers, de forgerons et de leveurs d’écorces, sans compter les chasseurs, braconniers et simples usagers. Peuplée de jour comme de nuit, théâtre d’une activité intense, la forêt était largement exploitée...
Si la Franche-Comté est aujourd’hui couverte à plus de 40 % de résineux et de feuillus en tout genre, on sait de manière certaine que ce taux était largement inférieur entre la fin du Moyen-Âge et le XVIIIe siècle. Ici comme ailleurs, la déforestation devient massive à partir du XVe siècle pour répondre à la fois aux besoins en énergie et en matériau de construction.
« Il ne faut pas oublier que la région est riche d’une tradition industrielle pluriséculaire, rappelle Paul Delsalle, historien à l’université de Franche-Comté, et que dès le Moyen-Âge, les usines comptent parfois jusqu’à deux ou trois cents ouvriers ! » Les salines disséminées sur tout le territoire en sont des exemples. Avec près de mille ouvriers au XVIIe siècle, celle de Salins-les-Bains (39) est sans conteste la plus importante. Trois mille cinq cents hommes s’emploient à la fournir en bois régulièrement ! L’exploitation de la forêt est soumise à des contraintes et des règles précises, cela des siècles avant le rattachement du Comté au royaume de France en 1678 et l’adoption des règles édictées par Colbert en matière de gestion forestière.
Certaines essences étaient réservées à l’industrie, et le droit des habitants se limitait en général au « mort bois » comprenant tilleul, noisetier et charme. Il était interdit de se servir en fruitiers, qui, outre les pommiers, poiriers, pruniers et cerisiers qui abondaient en forêt, comptaient aussi le chêne et le hêtre. Les dossiers de justice fourmillent de condamnations comme celle, au XVIe siècle, de ce Bisontin de retour de Chailluz, arrêté porte de Battant avec un chariot chargé de branches de cerisier. « Mais est-ce que nous ne surestimons pas la présence de ces variétés du fait qu’elles sont régulièrement citées dans les archives ? », se demande Paul Delsalle, qui voit d’un bon œil l’apport d’autres disciplines pour compléter les sources documentaires.

Archives et vestiges
Les travaux d’Aurore Dupin menés en forêt de Chailluz sont de cet acabit. Doctorante au laboratoire Chrono-environnement et rattachée à la MSHE où elle prépare une thèse en archéologie, la jeune chercheuse est spécialiste en anthracologie, l’étude des charbons de bois. Chailluz s’avère pour elle un excellent terrain d’investigation depuis que la technologie LIDAR (télédétection par laser aéroporté) a révélé les traces d’un millier de charbonnières, dédiées précisément à la fabrication du charbon, dont les résidus permettront d’identifier les essences d’origine.
« De nombreuses informations nous proviennent de la forêt de Chaux, où l’on perpétue encore la tradition du travail des charbonniers, explique Aurore Dupin. Pour la forêt de Chailluz, il n’existe plus de mémoire, peu de documents et pas de vestiges d’habitations qui toutes étaient construites en matériaux périssables. » Les méthodes scientifiques aident à pallier ce déficit. La susceptibilité magnétique confirme dans un premier temps les relevés du LIDAR. Elle certifie que l’argile du sol a subi des températures extrêmes. « Lorsque l’on chauffe fortement de l’argile, les minéraux qui la composent s’organisent d’une manière particulière, guidés par le champ magnétique terrestre. » La datation au carbone 14 atteste ensuite l’existence de la majeure partie des vestiges entre le XVIIe et le XIXe siècles. À partir d’infimes résidus, le microscope optique à réflexion est capable de déterminer l’essence du bois grâce à des caractéristiques anatomiques que le charbon présente sur trois faces. Paul Delsalle aura peut-être dans les mois qui viennent des réponses quant à la présence des fruitiers en forêt sur laquelle il s’interroge… Contact : Paul Delsalle - Aurore Dupin - Laboratoire Chrono-environnement - Université de Franche-Comté
Tél. (0033/0) 3 81 66 58 74 - paul.delsalle@univ-fcomte.fr / aurore.dupin@univ-fcomte.
La forêt, objet de convoitises
La difficulté à établir de façon précise des limites de propriété à l’intérieur des forêts n’est pas sans générer des tensions qui parfois tournent au pugilat. La forêt de Chailluz n’échappe pas à la règle et les Bisontins du XVIe siècle sont à couteaux tirés avec les habitants de Tallenay, de Chalezeule ou encore de Chatillon-le-Duc dans la défense de leurs lopins communaux. Une réalité d’autant plus criante que l’exploitation de la forêt est capitale à cette époque. Les habitants de Tallenay plantent même du Gamay sur les coteaux sylvestres en 1609. Mais la vigne s’avère difficile à entretenir, le vin de piètre qualité, et devant une production qu’il juge excessive, le Parlement de Dole ordonne l’arrachage des ceps. Le vin de Chailluz ne sera plus conservé que dans des pages d’archives...
Céline Bouvresse est enseignante en histoire et travaille régulièrement sur les forêts comtoises au travers de travaux de recherche universitaires. « Au XVIe siècle, les limites étaient fixées grâce à des points de repères naturels comme la crête d’une colline, ou d’autres plus discutables car potentiellement changeants : le tracé d’un chemin, la pose d’une borne en pierre ou la gravure d’un emblème sur un arbre. Les descriptions n’étaient qu’orales et on apprenait aux enfants à reconnaître les lieux. Il n’est pas rare que les dossiers de justice s’appuient sur les témoignages des anciens du village faisant appel à leurs souvenirs d’enfance pour servir de preuve. » Il faudra attendre le début du XVIIIe siècle pour que les premiers plans apparaissent et limitent les conflits en même temps que les propriétés.
Contact : Céline Bouvresse - Tél. (0033/0) 6 83 24 90 78 - bouvressec@yahoo.com
09:18 Publié dans Botanique, Environnement-Écologie, Paléontologie - archéologie | Lien permanent | Commentaires (0) | Facebook | | |
19/08/2014
Cactus géant
Cactus géant
par Dominique Delfino
photographe animalier, naturaliste et… curieux
Dans les ruelles du petit port du village de Bol sur l’île de Brac en Croatie je découvre un superbe Cereus de taille impressionnante grimpant le long d'un mur.
En opposition avec le chéneau qui recueille l'eau de pluie, la plante grasse s'élance vers le ciel, contourne la maison jusqu'à rejoindre le toit et la lumière qui lui profite tant.
Destin contraire de ces deux "organismes". Liés l'un à l'autre, si le chéneau se gorge d'eau de temps à autre, le cactus se gorge de soleil pour se développer sans que rien ne semble pouvoir l’arrêter.

Cliché © Dominique Delfino
08:47 Publié dans Botanique, Photographes naturalistes et scientifiques | Tags : dominique delfino, photographie naturaliste | Lien permanent | Commentaires (0) | Facebook | | |
20/07/2014
La forêt de Thise et sa gestion
La forêt de Thise
et sa gestion
par André Guyard
(dernière mise à jour :
19 juin 2018)
Cet article tire ses informations d'exposés et d'explications sur le terrain de MM. Joachim Hatton, ingénieur ONF et Daniel Moyne agent local ONF de Franche-Comté lors d'une intervention le samedi 22 juin 2014 dans le cadre du comité communal "Environnement".
La forêt de Thise a été jusqu'à la décennie 1950 la seule source de combustible pour le chauffage et l'industrie. Elle fut longtemps exploitée par des travailleurs spécialisés vivant en forêt avec leurs familles : charbonniers et bûcherons.
Les charbonniers fabriquaient du charbon de bois pour l'industrie ; c'était un combustible de qualité, donnant beaucoup de chaleur pendant longtemps. Désormais, les communes se tournent vers la production de bois d'œuvre qui constitue une ressource non négligeable.
Historique
Le lieu-dit la Gruerie à Thise évoque l'administration forestière d'avant la conquête française. Colbert ensuite, par son ordonnance des Eaux et Forêts de 1669, supprime les grueries et réglemente sévèrement l'utilisation de la forêt. Ces exigences sont mal acceptées par les utilisateurs ; ils poursuivent les anciens usages jusqu'à la fin du XVIIIe siècle comme par exemple mener paître les troupeaux dans les forêts avec un risque de surexploitation. Depuis l'ordonnance royale de 1827 qui constitue une grande réformation des forêts voulue par Colbert et qui est une reprise en main des forêts par l'État, celles-ci sont soumises à un contrôle plus sévère et elles sont aujourd’hui mieux gérées. Depuis 1964, c'est l'Office National des Forêts qui propose la gestion des forêts aux communes, ce qui est une bonne chose car on entre dans une nouvelle période où il y a une pression de plus en plus forte sur la ressource.
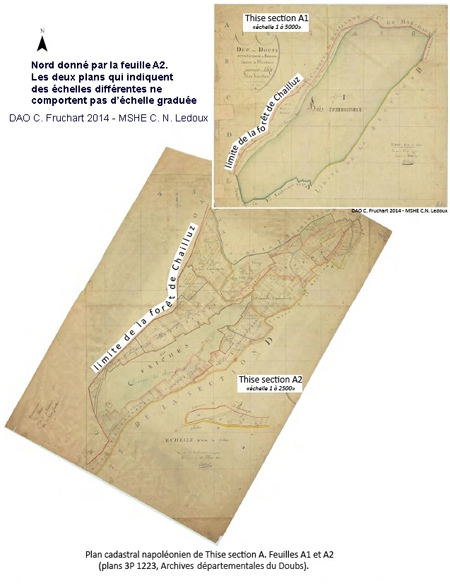
Une étendue importante
Notre bourgade côtoie un ensemble forestier compact de plus de 4000 hectares englobant le massif de Chailluz et son propre massif. La forêt de Thise s'étend sur 446 hectares 06 ares, ce qui représente plus de 47 % de la surface communale.
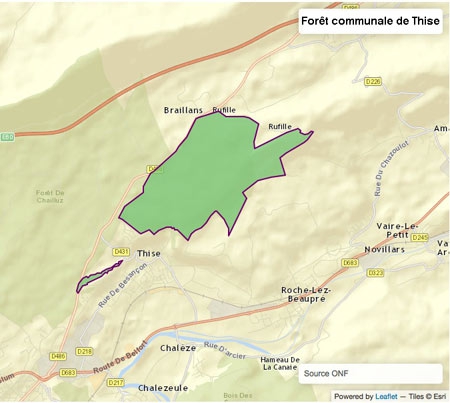
La forêt communale de Thise
Cette superficie forestière a évolué au cours du temps.
- 1804 : 263,20 hectares.
- 1838 : 383 hectares.
- De 1899 à 1920, les Communaux du Chemin du Roi l'agrandissent de 17,50 ha.
- De 1966 à 1970 il y a diminution de près de 10 ha suite à la construction des lotissements.
- En 1999, la commune se rend acquéreur de 55 ha au Bois du Fays sur la commune d'Amagney.
Situation géographique
Cette forêt est constituée par un massif principal d'accès facile qui occupe au Nord—Nord-Est du village un plateau à relief peu accusé, parsemé malgré tout de nombreuses dépressions (dolines) plus ou moins profondes (voir également sur ce même blog l'article sur le rôle de dolines dans l'érosion des sols de Franche-Comté). Il est en outre traversé par une légère dépression allongée du Sud-Ouest au Nord-Est, à l'Est de la grande sommière ; elle est marquée par une série de trous profonds dus à des effondrements. Cette dépression correspond à un cheminement souterrain d'une eau qui alimente la source du Paret. Le deuxième massif d'une surface de 6 hectares, occupe le versant Nord du canton de "La Côte des Buis".
L'altitude varie de 310 mètres au Sud du canton du "Coutelot et Grand Cotard", à 420 mètres au Nord du canton de "la Gruerie", à proximité de la ferme de Rufille. La forêt de Thise jouit d'un climat relativement doux. Une moyenne établie sur 76 ans donne une température moyenne annuelle de 10°1 et des précipitations égales à 1 100 mm par an.
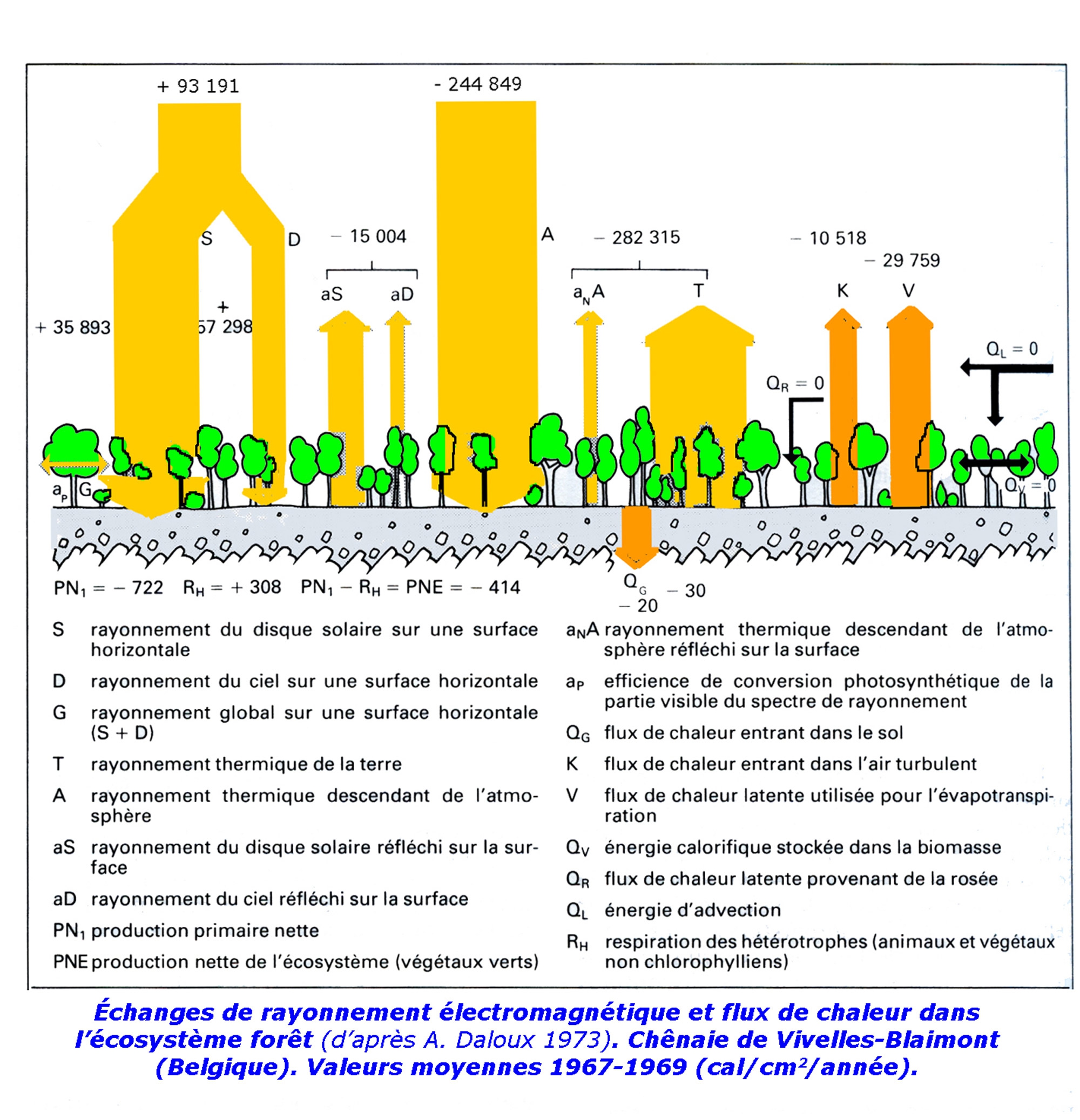
D'une façon générale, la forêt constitue un écosystème complexe où le climat et le sol ont une grande importance (voir l'article qui paraîtra prochainement dans ce même blog : la forêt de Chailluz). Le schéma ci-dessous montre l'importance des flux d'énergie qui traversent l'écosystème forêt. Pour plus de détails, on pourra consulter en ligne le cours de Jean-Yves Massenet ou l'ouvrage[1] de Sylvain Gaudin : Quelques éléments d'écologie utiles au forestier.
[1] édité dans le cadre du BTSA Gestion forestière Module D41 du CFPPA/CFAA de Châteaufarine (Besançon Doubs)
L'assise géologique
L'assise géologique est essentiellement calcaire. En série subhorizontale, on rencontre le Bajocien, le Bathonien, le Callovien, l'Oxfordien et le Rauracien.
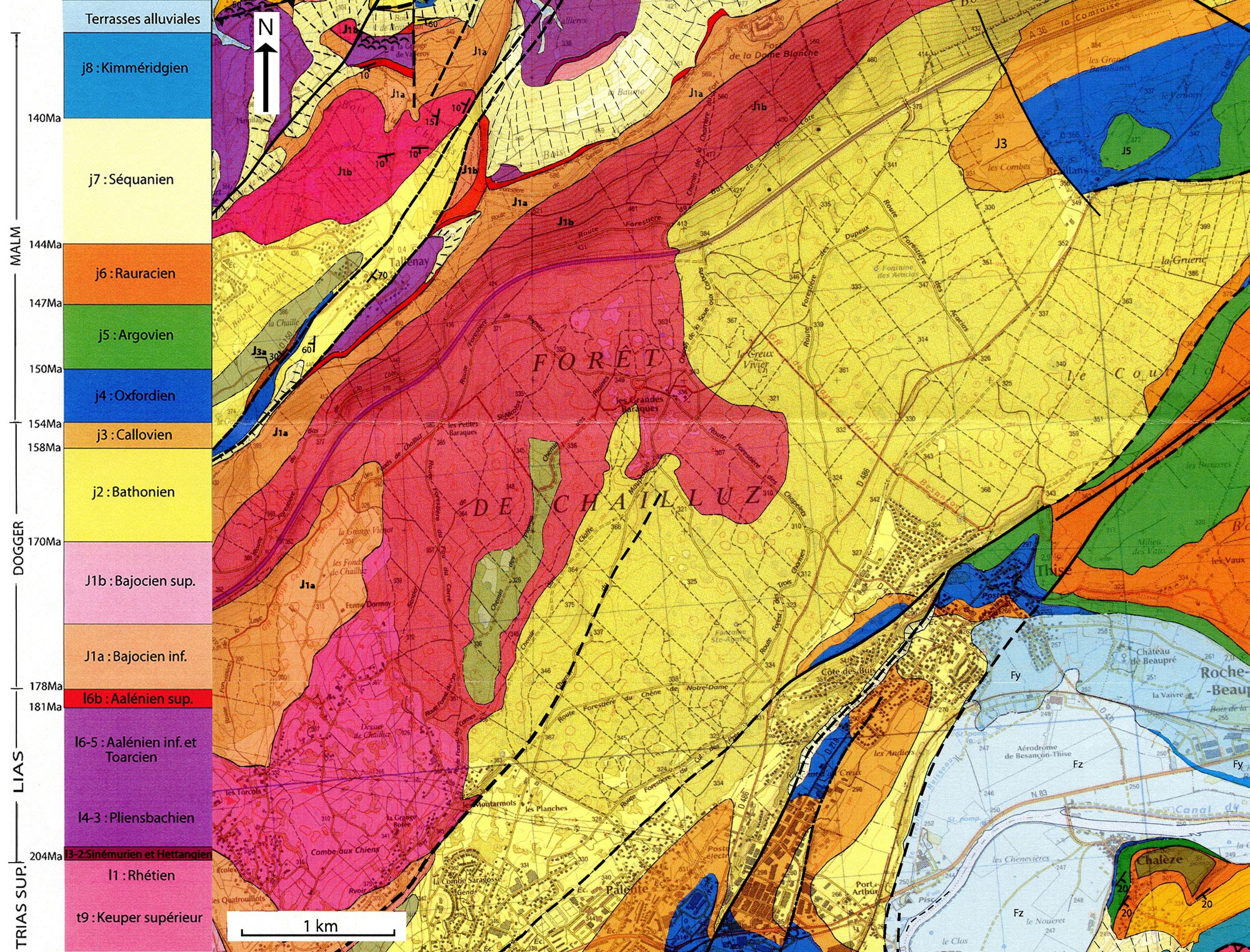
Assise géologique de la forêt de Chailluz-Thise
(d'après P. Rolin)
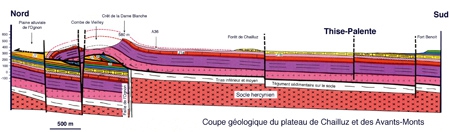
Coupe géologique du plateau de Chailluz et des Avants-Monts
(d'après P. Rolin)
Le sol
Avec le climat, la nature du sol constitue l'un des deux facteurs essentiels qui régissent la fertilité du sol. Sans renseignements précis sur les sols forestiers de la commune de Thise, le lecteur pourra consulter l'article dédié aux sols de la forêt de Chailluz dans ce même blog.
On peut se permettre de dire que les sols forestiers de Thise sont en général de bons sols plus ou moins évolués. Leur fertilité dépend de leur épaisseur variable selon les endroits et de l'exploitation.
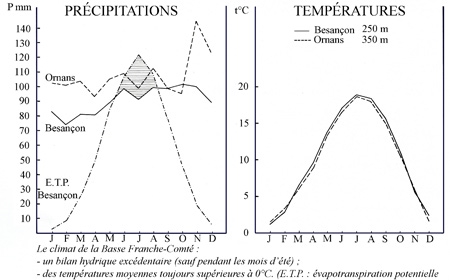
Aperçu du climat de la Basse Franche-Comté
Les principales essences forestières
Nous n'aborderons pas ici la flore herbacée qui se développe sous l'ombrage des arbres, ni des champignons que les mycologues et surtout mycophages débusquent dans les sous-bois. La flore de la forêt de Thise est en général calcicole. Toutefois, certains îlots ou placages siliceux, de décalcification ou de transport, permettent à la flore calcifuge de se développer.
La répartition des essences arborées comporte 80% de feuillus (hêtre principalement, chêne, frêne, merisier, érable, charme,) et 20 % de résineux (épicéa, sapin, pin).
Deux essences sont dominantes :
Le chêne (23 % en nombre) peut atteindre un diamètre de 0,60 m à 1,30 m du sol vers l'âge de 130 ans. Il est de qualité variable selon la fertilité de la station qu'il occupe.
Le hêtre ou foyard (20 %) atteint couramment un diamètre de 0,60 m à 10 ans. Il donne un bois tendre et blanc de bonne qualité qui se vend au cours le plus élevé pratiqué dans la région.
Le charme, constitue principalement le taillis, essence dominée, quelquefois réservé en futaie, il n'atteint jamais un gros diamètre et il est dominé par les autres essences et rejeté vers les stations les plus pauvres. Charme, tilleul et érable champêtre représentent 42 % des essences.
Parmi les essences disséminées, on rencontre le frêne, les érables (érable sycomore, érable plane, érable champêtre), les alisiers (blanc et torminal), le merisier, le bouleau, le tremble, le tilleul et le robinier faux-acacia (appelé acacia dans notre région) (7 %).
Parmi les résineux, on trouve quelques bouquets de pins et épicéas à l'Ouest de la forêt (7 %). Le sapin a été introduit dès 1927 dans le canton "La Gruerie". Il forme actuellement une futaie qui fournira des produits appréciés (poteaux, sciage). Son introduction devra néanmoins rester très localisée.
Ajout de janvier 2015 : la croissance du hêtre et de l'épicéa s'accélère en Europe depuis cinquante ans.
En 2014, une équipe allemande a pu montrer que la croissance du hêtre et de l'épicéa s'accélère en Europe depuis cinquante ans. Pour arriver à cette conclusion, Hans Preztech et ses collègues de l'université technique de Munich se sont appuyés sur la plus longue série d'observations de parcelles expérimentales de forêts en Europe, série commencée en 1872. Ils ont ainsi constaté que la vitesse de croissance des hêtres avait augmenté de 77 %, et celles des épicéas de 32 % par rapport à leurs niveaux de 1960. Plusieurs facteurs avancés : l'élévation des températures, rallongement de la période de croissance (le nombre de jours dans l'année dont la température dépasse 10°C), la hausse de la teneur en CO2 dans l'atmosphère et l'augmentation des dépôts azotés, qui ont tous deux un rôle fertilisant.
Attention cependant : une équipe franco-allemande a montré que la reforestation avec les conifères favorise le réchauffement du climat. La forêt européenne a gagné 1 200 000 km2 depuis 1750 avec le changement du mode de chauffage et le bond de la productivité de l'agriculture, qui ont provoqué le reboisement d'énormes surfaces agricoles. Malgré les vertus climatiques régulièrement attribuées à la reforestation, cette situation s'est en fait traduite, selon une modélisation publiée par cette équipe, par un réchauffement local d'environ 0,12°C (modeste, mais pas négligeable). "Depuis un siècle, beaucoup de conifères ont été plantés, qui se sont substitués aux feuillus", indique Aude Valade, de l'Institut Pierre-Simon Laplace, cosignataire de l'étude. Plus sombres, ils absorbent plus de rayonnement solaire. Un phénomène surtout marqué dans les zones enneigées où les forêts de feuillus et les champs, blancs tout l'hiver, sont très froids, et se réchauffent nettement une fois plantés de conifères." Sans compter que les feuillus transpirent plus de vapeur, et sont donc "rafraîchissants". "Reboiser ne suffit pas, conclut Aude Valade. Il faut tenir compte des essences et de la façon d'exploiter les parcelles." Plus de détails ici.

Forêt d'épicéas dans le canton de la Gruerie (parcelle 41)
Document © André Guyard
Généralités sur l'économie forestière et le traitement des forêts
II n'est point de fonction de l'écosystème sylvestre qui n'intéresse l'Homme. La réduction primaire nette (toutes matières végétales) ainsi que la production secondaire nette (gibier) ont été les plus exploitées. La récolte du bois s'est progressivement accompagnée d'une technique de régénération des arbres qui est devenue la sylviculture ; la chasse a évolué vers la cynégétique.
Dans les pays industrialisés, l'homme infléchit certains processus de l'écosystème sylvestre afin d'en tirer le maximum d'avantages ; il lui fait donc subir un traitement. Le traitement est l'ensemble des opérations que l'on pratique dans le but d'obtenir de la forêt, de façon soutenue, les services les plus adéquats ; production de bois, de gibier, de fruits, protection du sol et des eaux, loisirs, utilités sociales. Le mode de régénération caractérise le régime (taillis, taillis sous futaie, futaie).
Le traitement comprend deux séries d'opérations bien distinctes : la régénération (coupes, ensemencement naturel, plantation) et les soins culturaux (dégagement des espèces nobles de la compétition des espèces non économiques, élagages, éclaircies). Parmi ces derniers, l'éclaircie est l'opération la plus importante ; elle n'influence pas la production totale, mais modifie favorablement le diamètre des arbres. Elle permet de produire des fûts de grosses dimensions en un temps court. Les éclaircies se font à des intervalles plus ou moins réguliers dans le même peuplement ; c'est la rotation (de 4 à 8 ans).
On distingue ainsi la futaie, régénérée naturellement par les semences ou artificiellement par semis ou plantation, le taillis, qui après coupe se rajeunit naturellement par rejets de souches ou par drageons (chêne, charme, érable, frêne, bouleau), le taillis sous futaie ou taillis composé, qui comprend un taillis surmonté d'une strate arborescente d'espèces nobles (chêne, frêne, hêtre) se régénérant par semences (futaie) À l'heure actuelle, les régimes du taillis et du taillis sous futaie sont abandonnés, car ils ne répondent plus aux besoins de l'économie contemporaine.
Quelle que soit la forme des peuplements — régulière (peuplement équienne) ou irrégulière (peuplement d'âges multiples), les arbres se répartissent en classes sociales : dominants, codominants, intermédiaires, dominés. Les peuplements d'âges multiples sont constitués d'arbres d'âges variés, donc de tailles diverses. Les peuplements équiennes sont composés d'individus de même âge ; ils sont le plus souvent artificiels et réguliers (plantations d'épicéa, de pin).
Dans ces types de peuplement, on distingue le fourré, constitué de jeunes sujets dont les branches voisines se rejoignent et forment massif : le gaulis, constitué de gaules de moins de 10 cm de diamètre à 1,30 m au-dessus du sol ; le perchis, composé de perches de plus de 10 cm de diamètre ; la futaie, lorsque les arbres ont plus de 20 cm de diamètre et ont à peu près leur forme définitive.
La régénération d'une futaie équienne peut se faire au moyen d'une coupe unique (blanc étoc) suivie de plantation, par exemple pour l'épicéa. On peut également procéder à la régénération par la méthode des coupes progressives comportant : une coupe d'ensemencement qui desserre les cimes pour favoriser la fructification ; des coupes secondaires qui réduisent le couvert de la futaie au-dessus des jeunes semis et en favorise la croissance ; une coupe définitive qui enlève les derniers arbres de la futaie qui ont donné la semence, lorsque toute la surface est régénérée. Cette technique de rajeunissement, qui s'étend sur 20 à 50 ans, convient bien à la sapinière et à la hêtraie.
La révolution est le temps écoulé entre la naissance et la coupe des arbres mûrs ; elle est d'environ 60 à 100 ans pour l'épicéa, de 150 ans pour le hêtre, de 180 ans et plus pour le chêne.
La futaie jardinée est une forêt d'âges multiples présentant des arbres de tous âges et de toutes dimensions confusément mélangés (sapinière des Vosges, pessière du Jura). Elle est irrégulière et particulièrement esthétique. Le traitement consiste à parcourir la surface totale de la forêt et à enlever, ça et là, des sujets exploitables, soit pour éclaircir, soit pour régénérer. La régénération est permanente par petits bouquets. La futaie jardinée normale (en équilibre) comprend une gradation harmonieuse des classes de dimensions.

Flux d'énergie à travers les réseaux trophiques de la hêtraie jardinée
L'aménagement des forêts consiste en la réglementation des opérations culturales et d'exploitation en vue de donner au bénéficiaire un revenu annuel soutenu. En général, la superficie de la propriété forestière est divisée en un certain nombre de coupes (secteurs de forêt). Chaque année, on coupe ou l'on récolte dans une ou plusieurs coupes selon un plan à long terme (plan d'aménagement, règlement d'exploitation).En raison de la longévité des arbres et des arbustes, la masse végétale ou biomasse d'un habitat forestier est élevée. La valeur de la production ligneuse fluctue en fonction de son accroissement et des pertes qu'elle subit.
Matériel sur pied
La biomasse des hêtraies et des chênaies d'Europe est de l'ordre de 500 à 1000 tonnes à l'hectare. (Elle peut atteindre de 4000 à 6000 tonnes à l'hectare dans la forêt montagnarde du Jura, des Alpes, de Bohême à Picea excelsa et Abies alba, conifères qui peuvent attendre 50 m).
En économie forestière, on ne considère que le matériel « fûts sur pieds », soit la partie la plus utile à l'homme. Ce matériel est alors représenté par des valeurs plus modestes évaluées en mètres cubes. Le tableau 5 rassemble quelques données pour des jeunes peuplements créés artificiellement en Europe, où l'on voit que la masse peut approcher 800 m3. Des volumes exceptionnels se trouvent au Japon dans un peuplement de Cryptomeria japonica de 139 ans (2 806 m3), et sur la côte pacifico-américaine dans un massif de douglas de 87 m de hauteur (3 695 m3).

Population de douglas dans la parcelle 50 de Thise
Document © André Guyard
Productivité primaire et production de bois utile
Les végétaux croissent et se développent en relation avec les processus de photosynthèse ; la biomasse n'est donc pas rigoureusement stable. D'une part, elle s'accroît de nouvelles matières sous la forme de tissus et d'organes ; d'autre part, elle subit dans le même temps des pertes par mortalité d'organes : les houppiers accroissent leur sommet, mais en même temps perdent des rameaux et des branches à leur base ; des arbres naissent, tandis que d'autres dépérissent. La productivité primaire nette, processus cumulatif irréversible, comprend donc, en principe, non seulement la différence des biomasses entre deux temps donnés, gain restant acquis à la structure, mais également la fraction caduque ou prélevée (feuilles, rameaux, individus dépéris, récoltes, consommation par herbivores). La production primaire nette des forêts peut ainsi varier de 4 à 30 tonnes de matières sèches par hectare et par an. Les chênaies d'Europe occidentale produisent environ 12 tonnes (P. Duvigneaud, 1980).
L'économie forestière actuelle s'intéresse presque exclusivement à la productivité en bois de fût, soit une petite fraction de la productivité primaire nette (2 tonnes de bois de fût sur 12 tonnes de matières sèches totales.
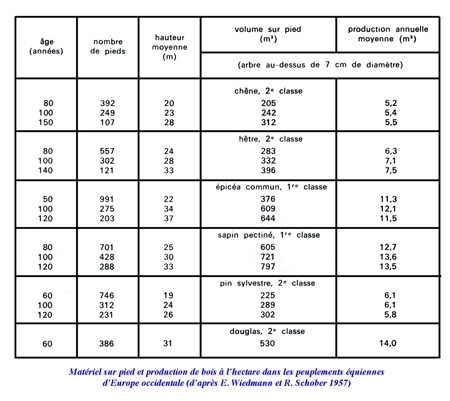
Le tableau ci-dessus donne les productions en bois de fût d'un certain nombre de peuplements équiennes européens. Elles varient de 5 à 14 m3 par hectare et par an. Ce sont les feuillus à bois dense (densité : 0,70) qui produisent le moins de volume, le hêtre se montrant supérieur au chêne. Les résineux à bois dense (pin sylvestre) produisent un volume voisin de celui des feuillus. Par contre, les conifères à bois léger et à fût se prolongeant haut dans le houppier (densité : de 0,45 à 0,50) produisent de 11 à 14 m3. Le douglas, espèce nord-américaine, donne en Europe des productions supérieures aux essences européennes.
Tous les peuplements d'une essence donnée ne produisent pas exactement les quantités de bois indiquées dans ce tableau. La production varie avec les sites et notamment avec la fertilité du sol. C'est la raison pour laquelle on établit des classes décroissantes de productivité. Ainsi, les hêtraies belges se répartissent sur cinq classes (de 1 à V), lesquelles correspondent à des groupements végétaux déterminés : la hêtraie à aspérule se situe dans la classe 1 avec un accroissement annuel moyen de 8,8 m3 ; les hêtraies à myrtille et à Calamagrostis dans les classes IV et V avec un accroissement de 2,8 m3.
La gestion de la forêt communale de Thise
La production de bois d'œuvre
L'effondrement des cours du bois de chauffage suite aux années cinquante incite la commune à se tourner vers la production de bois d'œuvre qui permet de tirer des revenus importants ; même à l'ère du béton armé et du plastique, le bois reste un matériau de première nécessité et beaucoup reviennent à son utilisation traditionnelle. L'industrie a besoin de bois d'œuvre en quantités de plus en plus importantes... Il est clair que la forêt doit être aménagée pour répondre aux besoins.
La coupe doit être vidangée pour une date fixe. Les arbres sont alors abattus, le bois d'œuvre mis en grumes et débardés par des bûcherons professionnels. Quant aux houppiers et aux arbres de moindre valeur, ils sont réservés pour l'affouage.
La coupe affouagère
L'affouage est une vieille coutume, un droit d'usage de la forêt par les habitants. À Thise, l'affouage a été abandonné vers les années 1970 en raison de l'augmentation du nombre d'habitants et surtout le peu d'amateurs pour la hache et la scie. Mais depuis quelques années, avec l'enchérissement du fuel, une cinquantaine d'affouagistes se déclarent en mairie et se partagent des lots d'une importance de 30 stères.
Chaque année une estimation de la coupe affouagère est assurée par l'Ingénieur des Forêts et autorisée par le Préfet : au total, une trentaine de parcelles à délivrer à raison d'une par année.
La coupe est partagée en deux catégories de valeur différente :
— la demi-portion représentée par les taillis et branchages qui donnent la charbonnette et les fagots ;
— la demi-portion constituée par le beau bois, plus apprécié car il "tient" le feu. Elle comprend les "modernes" de 25 ans d'âge, chablis cassés par la tempête ou arbres jugés sans avenir. Ces arbres sont numérotés sur la souche et le tronc à la peinture ou martelés au moyen d'un marteau spécial.
Ces deux demi-portions sont réparties en lots estimés par des garants et tirés au sort et partagées entre les villageois qui se déclarent en mairie.
Le rôle de l'ONF et l'Aménagement forestier
La gestion de la forêt communale est assurée par l'Office National des Forêts (ONF) par le biais d'un Aménagement forestier, c'est-à-dire un plan de gestion rigoureux qui a débuté depuis 1964. Ce document est un plan de gestion d'une durée de 20 ans dont le dernier renouvellement s'est effectué en 2012. Ainsi, l'Aménagement forestier en cours de la forêt communale de Thise se terminera en 2031.
L'Aménagement s'appuie sur la consolidation des aménagements passés, il en actualise les orientations stratégiques (poids relatif donné à la production, l'environnement, l'accueil du public). C'est le document cadre de la gestion de la forêt (art. L212-1 du Code forestier) pour une durée de 20 ans. Il est rédigé par l'ONF, validé par arrêté préfectoral et par délibération du conseil municipal. Le respect de l'aménagement garantit une gestion durable de la forêt. L'ONF et la DRAAF sont chargés de veiller à sa bonne application. Sa bonne observance donne droit à un label de certification PEFC (développement durable).
Rappelons que l'Office national des forêts (ONF) est un établissement public sous la tutelle de l'État. Il est l'unique gestionnaire chargé de la mise en œuvre du régime forestier dans les forêts de l'État et des collectivités (art. U21-3 et L111-1 du Code forestier).
Pour les forêts des collectivités, le régime forestier comprend notamment :
— l'élaboration d'un aménagement forestier en concertation avec le propriétaire ;
— la proposition de l'état d'assiette annuel des coupes, le marquage des bois et leur mise en vente ;
— la proposition du programme annuel de travaux ;
— la surveillance générale de la forêt.
Le financement de cette gestion est assuré :
— à 20 % par les collectivités propriétaires via les frais de garderie (12 % des recettes) et la taxe à l'hectare (2€/ha) ;
— à 80% par l'État via le versement compensateur.
Au-delà de la mise en œuvre du régime forestier, l'ONF réalise des prestations de service dans un cadre conventionnel (réalisation des travaux sylvicoles, maîtrise d'œuvre, assistance technique à donneur d'ordre).
Les choix techniques (essences, type de peuplement…) de ce document sont proposés à la commune par l'ONF, mais la commune participe à l'élaboration de l'Aménagement Forestier.
En application du régime forestier, la commune est propriétaire de la forêt. Elle décide des orientations stratégiques pour sa forêt, approuve l'Aménagement forestier, approuve le programme des coupes, décide du programme de travaux et accorde les concessions.
L'ONF assure la surveillance générale de la forêt communale (police forestière, chasse, nature), élabore l'aménagement, veille à son application, assure le marquage des bois, met en vente les bois, prépare les ventes, contrôle les exploitations.
L'ONF propose le programme annuel des travaux et veille à leur cohérence avec l'aménagement.
Parmi ces travaux, l'ONF procède à des cloisonnements, c'est-à-dire à des ouvertures au gyrobroyeur tous les 4-5 m de façon à permettre l'accès aux exploitants et affouagistes et par les forestiers pour les dégagements pour enlever des essences indésirables comme le charme qui gênent au développement des essences plus nobles comme le chêne et le hêtre. On recherche par là un peuplement de 100 arbres à l'ha.
Dans la forêt de Thise, le mode opératoire du traitement de la forêt qui était fondé sur la futaie régulière passe partiellement en mode de futaie irrégulière (comme le long du lotissement du Fronchot) pour favoriser l'aspect paysager. En futaie régulière, on procède à des coupes de régénération.
Programme de gestion spécifique de la forêt communale de Thise
— un diagnostic de la forêt est fait : potentialités, peuplements en place, desiderata de la commune. Partant de ce diagnostic sera établi un
— Programme d'exploitation.
Les objectifs de la gestion de la forêt de Thise sont multiples :
- produire du bois d'œuvre feuillus et résineux ;
- assurer le rôle écologique de la forêt en veillant à l'harmonie et à l'équilibre des différents étages de la chaîne alimentaire (production, consommation, prédation, décomposition) ;
- assurer l'accueil du public (fonction sociale) ;
- entretenir le paysage.
Pour atteindre ces objectifs, certaines zones sont dédiées à certaines fonctions.
• Fonctions de la forêt :
|
Fonctions principales |
Sans objet |
Enjeu faible |
Enjeu moyen |
Enjeu fort |
|
Production de bois |
3,36 ha |
18,90 ha |
211,70 ha |
211.65 ha |
|
Fonction écologique |
— |
445,56 ha |
— |
— |
|
Fonction sociale (accueil du public, paysage) |
— |
345,56 ha |
100 ha |
— |
En ce qui concerne la production de bois, la gestion utilise différents modes de traitement.
• Choix des modes de traitement :
1. Futaie régulière (405,82 ha). Sur la forêt de Thise on n'a pratiquement que des peuplements réguliers. Elle se caractérise par des peuplements homogènes avec phase de régénération périodique (80-160 ans).
Parcelle dont tous les arbres ont le même âge, la même hauteur et le même stade de maturité. On passe du stade semis au stade fourré, gaulis, jeune futaie, futaie mûre (parcelles 18-20) dans laquelle on va faire de la régénération. La régénération entraîne la coupe de tous les arbres (coupe blanche) pour revenir au point de départ : le semis. La chronologie de ce type de traitement s'établit selon le schéma ci-dessous :
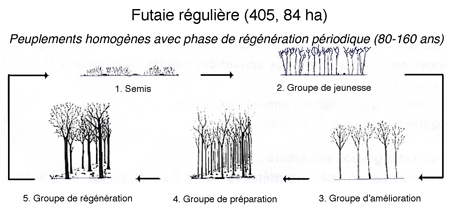
- Groupe de jeunesse (semis) : pas d'arbres adultes.

Groupe de jeunesse non éclairci (parcelle 40)
Document © André Guyard

Groupe de jeunesse éclairci (parcelle 41)
Document © André Guyard

Un cloisonnement ou layon dans la parcelle 40 permettra la pénétration des engins forestiers
Document © André Guyard
Entre ce groupe et le prochain, on peut récolter des baliveaux pas encore du bois d'œuvre (diamètre 20-25 cm) pour éclaircir le peuplement (affouage).
- Groupe d'amélioration : arbres déjà exploitables en partie, mais pas de phase de récolte. On retire les arbres en mauvais état (coupes sanitaires), les arbres les moins beaux pour permettre un meilleur développement aux autres (coupes d'amélioration = éclaircies), parcelles 41, 42, 43 = gaulis (20-35 cm de diamètre).
- Groupe de préparation : récolte sanitaire d'arbres dépérissants, équilibrer au niveau des essences les plus intéressantes afin de préparer la phase de régénération.
- Groupe de régénération : stade de récolte qui dure de 10-15 ans. Il y a étalement des récoltes entre les diverses parcelles. En 2015, seront récoltées les parcelles 30i, 31, 37 et 38. Parcelle de régénération : 33.

Groupe de régénération (parcelle 18)
Document © André Guyard
|
Groupe de jeunesse (semis) |
93,70 ha |
|
|
Groupe d'amélioration |
feuillus |
118,73 ha |
|
résineux |
48,40 ha |
|
|
Groupe de préparation |
72,31 ha |
|
|
Groupe de régénération |
72,68 ha |
|
Répartition des groupes en superficie
2. Futaie irrégulière (31,41 ha). Elle se caractérise par des peuplements hétérogènes avec processus de régénération en continu.
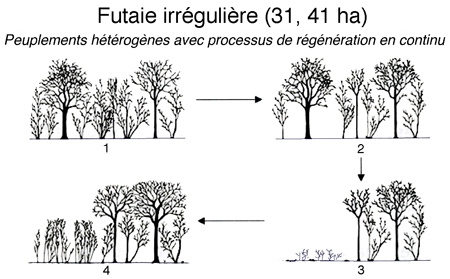
Pas de stade ouvert. Le semis de chêne est désavantagé, car c'est une essence qui réclame de la lumière. (parcelles 26, 30, 31, 32). Ce sont des parcelles périphériques gardées en paysager pour des questions esthétiques. La régénération est difficile.
|
Groupe irrégulier |
31,41 ha |
Répartition en superficie
La forêt de Thise comporte 57 parcelles. Un inventaire précis fixe tous les 20 ans, par parcelle, le contenu des différentes essences diamètre par diamètre. D'après cet état des lieux, une étude entreprise sur 20 ans définit la régénération d'un nombre donné de parcelles, c'est-à-dire le remplacement des peuplements vieillis ou trop pauvres par de jeunes semis ou des plantations. Les moyens financiers nécessaires pour mener à bien l'opération sont évalués ainsi que l'ordre de passage en coupe pour toutes les parcelles.
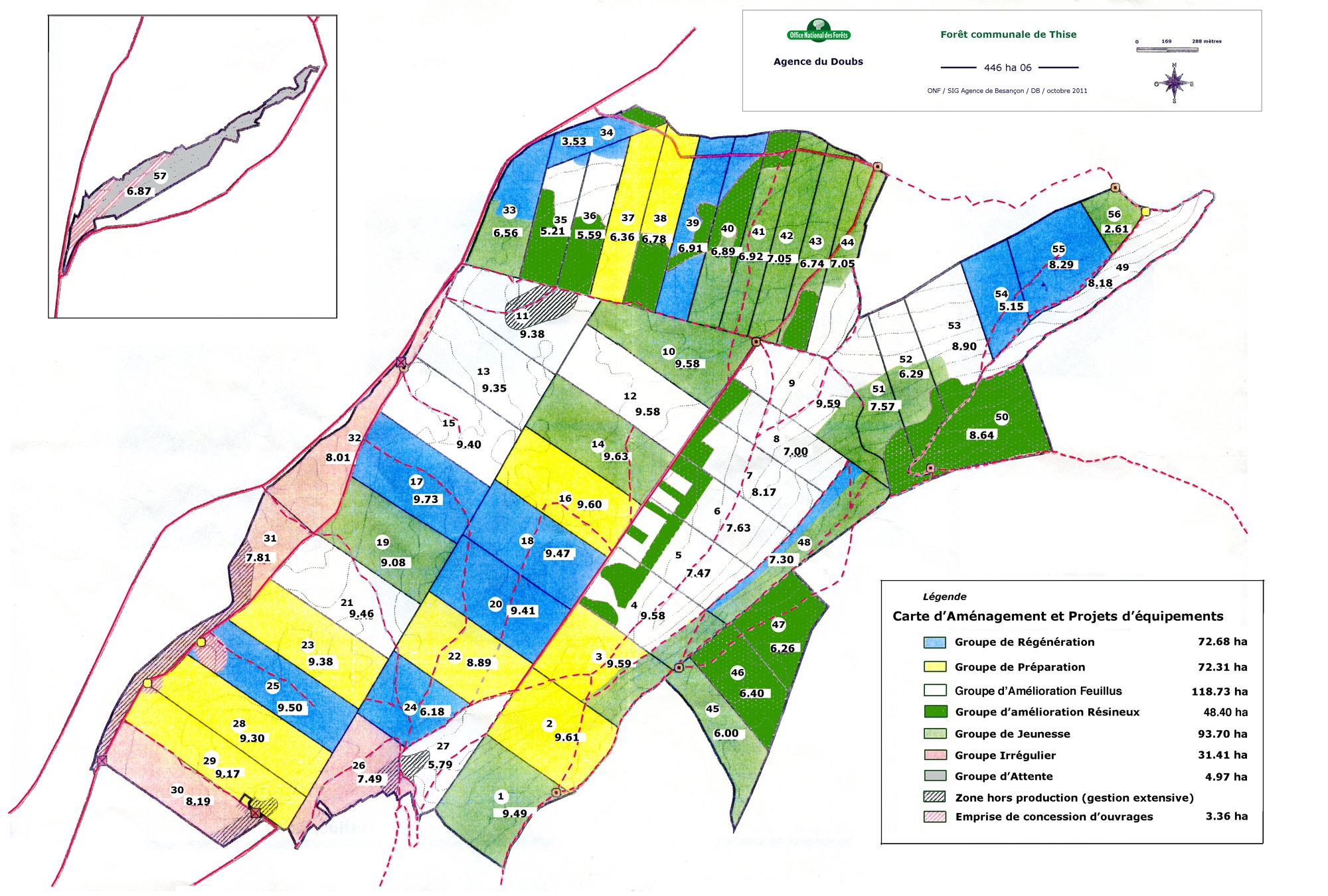
Répartition des 57 parcelles de la forêt de Thise
L'aménagement de taillis sous futaie ne répondant plus aux besoins humains, on s'oriente vers la production de bois d'œuvre. C'est l'objet de l'aménagement qui a débuté dès 1964.
Le programme actuel s'étend de 2012 et se poursuit jusqu'en 2032. Ainsi en rajeunissant le 1/6 de la forêt tous les 20 ans, on peut raisonnablement prévoir que les peuplements de la forêt de Thise seront renouvelés tous les 120 ans sauf pour la population de chênes renouvelée au bout de 160 ans.
En 2016, l'ONF annonçait qu'un essai de production de sapins de Noël serait tenté.
Cycles naturels des écosystèmes forestiers (Ajout du 19 juin 2018);
Divers travaux ont permis d'approcher de manière cyclique l'évolution des peuplements en forêt non exploitée. MAYER (1976) et LEIBUNDGUT (1982) ont proposé un modèle général qui retient la technologie suivante pour décrire ce cycle :
- Phase initiale : peuplement dense, ferme, dominé par les petits bois.
- Phase optimale : peuplement ferme à bois moyens dominants et à surface terrière élevée.
- Phase terminale (ou de sénescence) : peuplement encore relativement ferme, riche en gros bois avec peu de régénération.
- Phase de déclin : le peuplement s'ouvre avec des chablis. Le volume sur pied chute et la régénération apparaît.
- Phase de rajeunissement (ou de régénération) : phase ultime de la dégradation de l'ancien peuplement caractérisé par l'abondance de la régénération et des perches.
À ces 5 phases présumées s'enchaîner de manière cyclique, s'ajoute une sixième dite «jardinée», caractérisée par un équilibre des différentes catégories de hauteur et de diamètre des arbres. Cette phase est en général transitoire et résulte d'une phase de déclin-rajeunissement très étalée.
L'ensemble des phases s'organise dans le schéma théorique suivant :
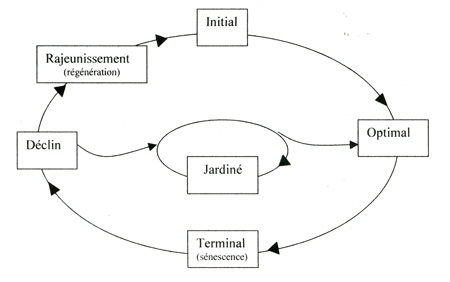
Dans le contexte européen de forêts à « dynamique douce », les unités de régénération sont plutôt de petite taille (15 à 50 m2). Dès qu'une perturbation génère l'ouverture d'une nouvelle unité, un nouveau cycle démarre. Ainsi, les nouveaux cycles commencent habituellement avant que les anciens ne soient totalement achevés. Plusieurs phases peuvent donc se chevaucher sur une même unité : la phase de régénération d'un nouveau cycle débutant dès que les premiers arbres morts d'un cycle ancien (en phase de sénescence) permettent à la lumière de percer la canopée.
- KORPEL (1995), dans son important travail sur les hêtraies-sapinières, complète les travaux de H. LEIBUNDGUT et organise ces 5 phases au sein de 3 stades successifs (fig. suivante) :
— le stade de régénération ou de dégénérescence, comprenant simultanément :
- la phase de sénescence constituée d'arbres mourants du cycle 1
- la phase de régénération constituée de jeunes semis du cycle 2
— le stade d'accroissement, comprenant simultanément :
- la phase de déclin constituée d'arbres morts du cycle 1
- la phase initiale constituée de jeunes arbres du cycle 2
— le stade optimal, constitué par la phase optimale représentée par des arbres en pleine croissance du cycle 2.
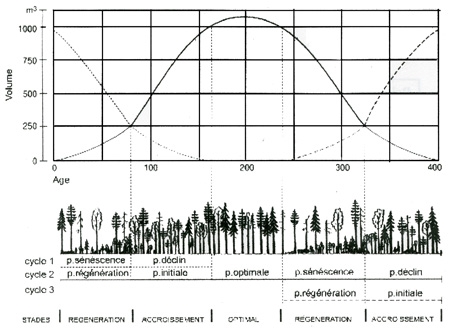
Modèle de sylvigenèse pour une hêtraie-sapinière à dynamique douce. Dans sa partie supérieure, la figure présente l'évolution de la biomasse (en m3/ha) pour chacun des trois cycles sylvigénétiques concernés : fin du cycle 1 (ligne pointillée), cycle 2 complet (ligne pleine) et début du cycle 3 (ligne entrecoupée). D'après S. KORPEL — 1995.
Ressources forestières de la commune
Chaque année la commune de Thise perçoit des revenus des différentes ventes (résineux et feuillus).
Une partie des recettes est investie dans des travaux forestiers (plantations de plants forestiers, travaux de dégagements de plants forestiers ou de semis naturels, travaux divers qui permettent à la forêt de se renouveler.
Le traitement des peuplements forestiers se fait sur la durée. Avec le réchauffement climatique, il est probable que d'ici 100 à 160 ans, on ait de fortes chances de procéder à des changements radicaux dans la nature des essences. On doit donc s'y préparer en favorisant des essences résistantes à la sécheresse et le chêne résiste mieux à la sécheresse que le hêtre. C'est une raison supplémentaire pour traiter la forêt en futaie régulière de chênes.
Les ventes de bois sont des ventes d'arbres sur pied désignés par martelage ou peinture dans les parcelles. Dans ce cas, l'abattage, le traitement et le débardement des grumes sont à la charge de l'acquéreur.
Mais l'ONF procède de plus en plus à la prévente du bois façonné. Dans ce dernier cas, les arbres sont façonnés par la commune avec l'assistance de l'ONF. Les bois se retrouvent en lots homogènes par essence au bord des routes prêts à être embarqués. Les prix de vente varient entre 80 et 200 € le m3 en fonction de la qualité.
Annuellement, les ressources communales issues de la vente des bois se montent à 40-41 000 € nets, déduction faite des programmes de travaux, des frais de garderie de l'ONF (12 %), ce qui paye 20 % de la gestion, les 80 % restants étant assurés par l'État. Depuis 2012, la FNCOFOR (Fédération Nationale des communes forestières de France) et l'ONF ont mis en place une taxe de 2€ à l'ha pour l'ensemble des communes forestières françaises. Les dépenses forestières ne représentent même pas 3 % des dépenses totales.
Les dégâts entraînés par l'exploitation et le débardage doivent être réparés par l'exploitant ou l'affouagiste sous risque de condamnation pénale.
La faune de la forêt de Thise
Il est rare d'apercevoir les mammifères hantant en liberté le massif forestier Chailluz-Thise. Ce sont essentiellement des rongeurs (campagnols, mulots, souris), des lièvres, des insectivores (taupes), des Ongulés (sangliers, chevreuils et même cerfs dans la forêt de Chailluz et chamois dans la côte de Bonnay), des mustélidés (belettes, fouines, hermines, blaireaux), renards, chats sylvestres.
En revanche, on peut rencontrer et écouter les chants de nombreux oiseaux : pinsons, merles, mésanges charbonnières, mésanges bleues, mésanges à tête noire, mésanges à longue queue, queues-rouges, rouge-gorge, pics épeiches, pivert, buses etc.
Il n'est pas rare de rencontrer des reptiles en particulier la Couleuvre verte et jaune. Les amphibiens se font plus discrets en raison de l'absence de collections d'eau pour la reproduction.
Accueil du public
La forêt communale est un espace ouvert à tous en respectant certaines règles. En premier lieu, le grand public recherche un espace arboré agréable et reposant, un air vivifiant et une atmosphère éloignée des bruits de la ville et de la circulation urbaine. Parmi les familiers de la forêt, les naturalistes qui consacrent leur vie à l'étude de la flore et de la faune sont enclins à protéger l'écosystème forestier. Les autres usagers : ramasseurs de champignons, chasseurs, pique-niqueurs, randonneurs ou promeneurs à pied ou en VTT sont susceptibles de fréquenter ce milieu avec respect !
Routes et chemins
L'accès aux différentes parcelles de la forêt doit être facilité par un réseau de routes et de chemins accessibles aux engins de traitement des bois et de débardage. Accessoirement, des sentiers peuvent être aménagés pour les randonneurs, les piétons et les VTT.
Chemins actuels
Au Nord, la forêt de Thise est longée sur 1500 mètres par l'actuelle RD 486, et traversée sur 2 000 mètres par l'ancienne route royale ou Chemin du Roi.
Une grande sommière empierrée partage le massif en deux. Elle est prolongée au Sud jusqu'au village par un chemin également empierré. Ces deux voies traversent la forêt thisienne sur 5 200 mètres. Un chemin traversant le canton "La Gruerie" relie Braillans à la ferme de "Rufille".
En 1975, la commune a apporté une amélioration sensible au réseau en élargissant et en goudronnant ce chemin et en le reliant à la grande sommière, par la création d'une route forestière goudronnée sur 0,700 km. Un autre chemin empierré permet d'accéder au Sud du canton "Le Grand Cotard". De plus, des chemins accessibles aux engins de débardage traversent les différents cantons.
En 2014, afin de faciliter le débardage des bois part les grumiers, un chemin empierré a été prolongé jusqu'au (40 % de subvention de l'État).
Chemins anciens
Le chemin du Paret, appelé chemin Perret sur d'anciens plans, se prolonge sur Braillans par le chemin dit "du Facteur" présentant un raccourci jusqu'au bureau de poste cantonal.
Le chemin du Fays qu'une erreur de lecture fait appeler le chemin du Fou court vers la nouvelle parcelle achetée en 1999.
À la suite du 8e Plan a eu lieu le renforcement de l'empierrement de la grande sommière, l'ancien chemin d'Amagney. En 2012, la Commune a réalisé un sentier botanique en boucle avec des panneaux pédagogiques permettant d'identifier différentes essences. March'en Thise, la section de randonneurs de l'Avenir de Thise a balisé un sentier de randonnées permettant de découvrir un ensemble de dolines. Récemment en 2014, a été réalisé l'empierrement d'un chemin de débardage des bois permettant aux camions grutiers de faire demi-tour à son extrémité.
La forêt dans l'éducation
La fête de l'Arbre rappelé plus haut est un symbole fort pour routes les générations, en 1927 comme en 1985, année de la plantation place de l'Amitié de deux arbres symboliques :
- par les enfants : l'arbre dit des Droits de L'Homme en mars : un érable sycomore ;
- par le Bourgmestre de Partenstein, la commune allemande jumelée avec Thise et le Maire de Thise : le chêne provenant du Spessart en Allemagne.
La Municipalité a organisé en mars 1980 une sortie "connaissance de la forêt" à l'intention des scolaires et des adultes avec le concours des forestiers.
Des sentiers de randonnée sont établis pour les promeneurs, sportifs ou pas :
G.R. 59 Grande randonnée de Pays ceinture de Besançon. Ce parcours passe au Nord-Est du village de Roche-lez-Beaupré à la forêt de Chailluz par le chemin des Vaux et du Paret.
Un chemin a été aménagé qui permet un accès facile et fleuri au monument de la Libération. Une table d'orientation permet d'appréhender le paysage de Thise et de ses environs.
En outre des Circuits V.T.T. (vélo tous terrains) prolongent les sentiers existant en forêt de Chailluz.
Un sentier botanique avec des panneaux indicateurs des essences rencontrées a été inauguré en 2013.
Les randonneurs de la section March'en Thise de l'Avenir de Thise ont tracé un sentier de 12 km dit le sentier des dolines dans la forêt de Thise.
La forêt doit avoir une présence constante à toutes les générations. Il s'agit de mieux la connaître, de s'efforcer constamment de la protéger, de l'améliorer ; il est de notre volonté de gérer cet héritage en conciliant écologie et économie.
Sources :
- Documents fournis par MM. Hatton et Moyne.
- Thise d'hier et d'aujourd'hui de MM. Henri Masson, Georges Perrin et Claude Proudhon.
- Ramade F. (1987). - Éléments d'écologie — écologie fondamentale.
La forêt face au réchauffement climatique
Deux mois après la fin de la COP21 à Paris, un article scientifique[1] paru dans la prestigieuse revue Science, jette un pavé dans la mare : certaines gestions des forêts en Europe n’ont pas été vertueuses pour le climat depuis 1750 ! Mais qu’on ne s’y trompe pas, cet article ne fait que confirmer que toutes les forêts n’ont pas la même capacité d’atténuer le changement climatique, ni tous les modes de gestion. Une preuve de plus que le rôle des forêts dans l’atténuation du changement climatique est plus complexe qu’on voudrait nous le faire croire et une invitation à réfléchir aux orientations futures de la politique forestière.
Pour le climat, mieux vaut beaucoup de forêts, et des futaies feuillues !
L’article rappelle les grandes évolutions de l’histoire des forêts européennes depuis 260 ans : une forte extension des surfaces forestières, une sylviculture favorisant les futaies (grands arbres utilisés pour des usages à long terme – charpente, etc.) plutôt que les taillis (plusieurs petits troncs poussant sur une même souche, destinés au bois de feu), la mise en gestion de forêts auparavant inexploitées, ainsi que la conversion de forêts de feuillus (chêne, hêtre…) en résineux (pins, épicéa…). Les deux premiers facteurs ont permis d’augmenter le stockage de carbone dans les forêts, compensant ainsi une part des émissions de CO2 responsables du changement climatique. A l’inverse, les deux derniers ont eu un effet contraire, contribuant à aggraver le dérèglement climatique : plus de résineux et plus d’exploitation, la fausse bonne solution pour le climat ! Et dans les produits bois ? Peu d’espoir de ce côté-là, le stockage de carbone est de trop courte durée pour compenser les émissions dues à l’exploitation et au changement d’essences.
Il est encore temps d’appliquer une politique vertueuse pour le climat
Réjouissons-nous ! En France, les essences feuillues occupent les deux tiers de la surface totale des forêts. Mais cela pourrait bien changer, puisque le Programme National de la Forêt et du Bois, en cours d’élaboration par l’État, promeut notamment la transformation de forêts feuillues en résineux et l’intensification de l’exploitation. Ces orientations visent à satisfaire les demandes de certaines filières industrielles de court terme, qui instrumentent le changement climatique pour justifier de couper les arbres toujours plus jeunes et de planter des résineux, demandés par les marchés. Pour Hervé Le Bouler, pilote Forêt de FNE : « Nous n’avons de cesse de contester ce raccourci que l’on voudrait imposer, et cet article renforce encore nos arguments. De plus, il est en cohérence avec le rapport établi par l’ONERC[2] et rendu public en 2015, qui prévenait déjà contre les fausses bonnes solutions, ce qu’on appelle la mal-adaptation, ainsi qu’avec d’autres publications scientifiques, toujours plus nombreuses. »
Julie Marsaud, coordinatrice Forêt de FNE, conclut : « L’intensification de l’exploitation et la transformation de nos forêts pour satisfaire l’industrie sont des impasses du point de vue du climat. Les solutions doivent être recherchées ailleurs, en luttant contre le fléau de la déforestation, en favorisant le stockage de carbone dans les arbres et les sols plus longtemps, en maintenant les essences feuillues et en augmentant la diversité au sein des écosystèmes forestiers. »
Pour Michel Dubromel, Vice-Président de FNE, « Deux mois après avoir réussi à obtenir un accord sur le Climat à Paris, la France ne doit pas relâcher son engagement. Comme toutes les autres activités, la forêt jouera un rôle important : celui du stockage à long terme du Carbone ».
[1] Naudts et al. Europe's forest management did not mitigate climate warming, Science 351, 597 (2016)
[2] ONERC. L’arbre et la forêt à l’épreuve d’un climat qui change. Rapport remis au Premier Ministre et au Parlement.
22:13 Publié dans Botanique, Environnement-Écologie, Géologie-hydrogéologie-Climatologie, Patrimoine franc-comtois et jurassien | Tags : thise, forêt de thise, forestologie, sylviculture, jura, hêtre, chêne, office national des forêts, écologie forestière | Lien permanent | Commentaires (0) | Facebook | | |
Le rôle des dolines dans l'érosion des sols du massif Chailluz-Thise
 Le rôle des dolines dans l'érosion des sols
Le rôle des dolines dans l'érosion des sols
du massif Chailluz-Thise
par André Guyard
L'érosion des sols est un phénomène général qui a partout suivi le passage de la hache et du feu. Chaque année, c'est par milliers d'hectares que des terres sont soustraites à l'agriculture, emportées par l'eau ou le vent. Les seules tornades de la dernière décennie ont arraché dans les plaines de l'Ouest des États-Unis des centaines de millions de tonnes de sols. Des phénomènes dont les vidéos fleurissent sur Internet !
En Franche-Comté, le processus est heureusement plus limité, mais les déforestations imprudentes, la topographie, la rudesse du climat ou le surpâturage sont autant de facteurs qui, agissant ensemble ou séparément, sont susceptibles d'entraîner la disparition totale de la couverture pédologique comme dans le Haut-Jura. En outre, nous sommes dans une région karstique. Le lapiaz de Loulle dans le Jura illustre bien cette violence potentielle de l'érosion.
Des zones sensibles se rencontrent également dans les bassins limono-sableux de la Saône et de l'Ognon. Dans ces secteurs, il n'est pas rare de voir le bas des champs recouvert par de la terre venue du haut. Parfois, ces matériaux d'apport doivent être réensemencés, les cultures s'y trouvant totalement enfouies à la suite d'une pluie violente.
Quand l'entraînement des terres est plus discret, il n'en est pas moins réel et d'autant plus inquiétant qu'il concerne en priorité les particules fines, les plus importantes pour le maintien de la fertilité.
Un premier exemple de l'exportation de ces matériaux fins est illustré par les graphiques ci-dessous.
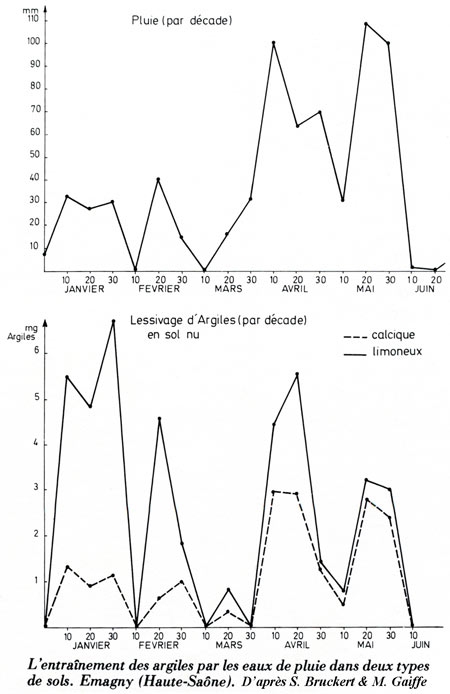
Il s'agit d'une évaluation des pertes en argiles par lessivage au cours du temps dans deux sols cultivés du secteur de Pin l'Émagny (Haute-Saône).
On remarque tout d'abord que l'intensité du lessivage, évolue parallèlement à l'abondance des précipitations.
Mais à l'intérieur de ce rythme climatique apparaît très nettement l'influence du type de pédogenèse : le sol calcique, dont les agrégats soudés par le calcium sont très stables, retient beaucoup mieux ses argiles que le sol limoneux dont la structure est plus fragile. Cette observation devrait nous rassurer sur la stabilité des sols des plateaux jurassiens, argilo-limoneux calciques et couverts presque en permanence par la forêt ou la prairie. Les travaux récents ont pourtant montré que même dans ces conditions favorables se manifestent d'importants mouvements de matériaux, affectant soit des sols entiers, soit des particules fines.
Dans les régions calcaires karstiques, les actions de dissolution des roches et les circulations d'eaux souterraines se traduisent en surface par des effondrements généralement connus sous le nom de dolines. C'est le cas du massif de Chailluz-Thise qui est criblé de ces entonnoirs. Des alignements de dolines signifient la présence d'une rivière souterraine dans le karst sous-jacent. La technique du lidar révèle de façon spectaculaire comment le sol de nos forêts est troué de ces formations.
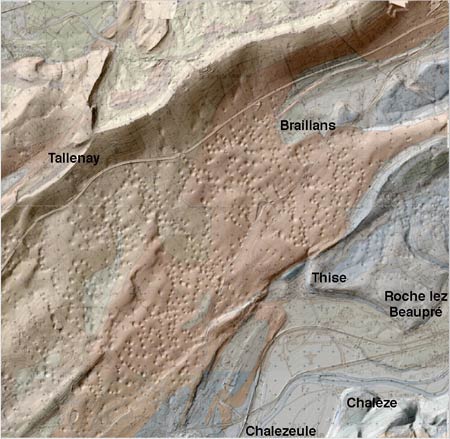
Image Lidar du massif forestier Chailluz-Thise
Qu'est-ce qu'une doline ?
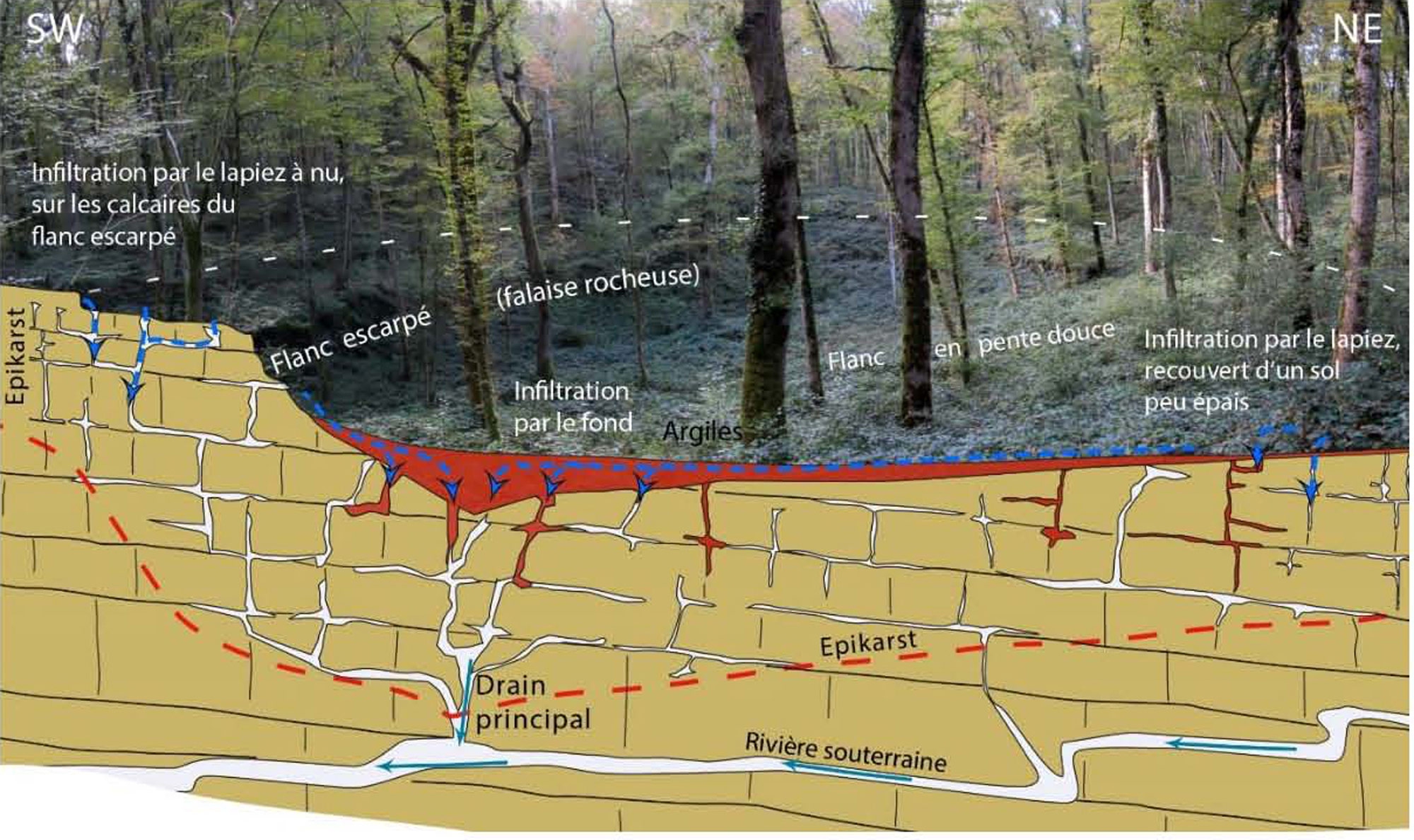
Coupe d'une doline dissymétrique de la forêt de Chailluz
Schéma © Patrick Rolin (cliquer pour agrandir le document)
Bizarrement, les dolines sont souvent dissymétriques avec un versant en faible pente du côté nord-est et un versant abrupt du côté sud-ouest. Le fond de ces dolines est relié aux galeries souterraines par des fissures ou des petits conduits verticaux de l'épikarst, souvent obstrués par les argiles. Le schéma ci-dessous explique comment cette dissymétrie peut se justifier.
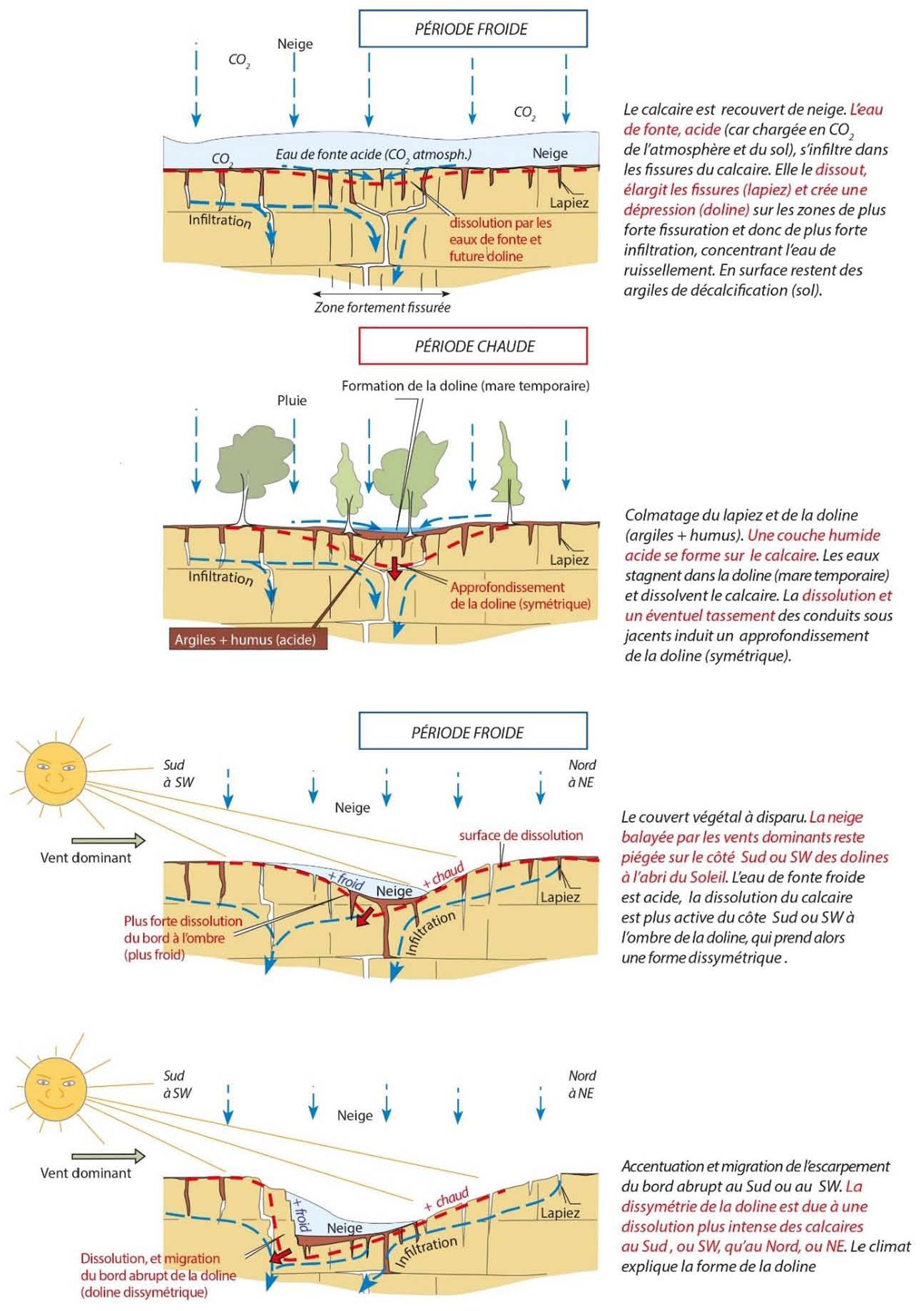
Évolution d'une doline Schéma © Patrick Rolin
(cliquer pour agrandir le document)
Les dolines se développent sur un substratum calcaire massif et épais, à partir de légères dépressions installées sur une zone fracturée, concentrant tes eaux de ruissellement. L'action dissolvante de l'eau de pluie est favorisée par son acidité due à la présence de gaz carbonique dissous. Cette eau en s'infiltrant dans les fractures et les joints des calcaires attaque la roche et creuse des cavités de forme variée. La dissymétrie d'une doline est due à une dissolution plus intense des calcaires à l'Ouest qu'à l'Est.
Au niveau des dolines, on a pu mettre en évidence des phénomènes extrêmement originaux, qui se traduisent par un triple mouvement : le remplissage des dolines, le lessivage de leurs sols et l'évacuation des matériaux par le fond.
Les dolines sont alimentées en matériaux par les ruissellements
Au fond des dolines, on retrouve en coupe le sol ancien recouvert progressivement par les apports de limons dus au ruissellement.

Sol de doline. L'ancienne surface du sol a été enterrée
sous les apports © S. Bruckert
Parfois se manifestent des anomalies dans ces dépôts en milieu karstique. Par exemple, dans une doline du massif du Risol, le sol brun jaunâtre profond présentait, à environ vingt centimètres de la surface, une bande grisâtre qui ne pouvait pas s'expliquer par un phénomène pédogénétique.
Des lames minces effectuées à ce niveau ont permis de reconnaître de nombreux débris organiques peu transformés, en particulier des tiges et des feuilles de graminées. Des figures identiques se retrouvaient dans les premiers centimètres du sol, permettant d'assimiler les deux niveaux. La bande grisâtre représente donc un horizon atypique, c'est-à-dire la surface d'un ancien sol, enfouie sous des sédiments plus récents dans lesquels s'est réimplantée la végétation actuelle.
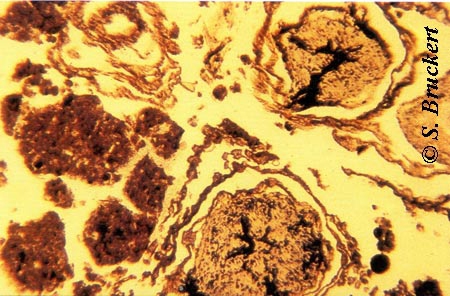
Dans l'ancien horizon de surface subsistent des traces de l'ancienne végétation
(tiges et feuilles de graminées mélangées aux limons)
On peut également trouver dans le fond des dolines des traces des activités humaines. La coutume millénaire qui incite les bûcherons à allumer leurs feux dans les bas-fonds clairières et bien protégés du vent, a permis une observation intéressante dans une doline de Bonnevaux (La Vieille Citerne). Le sol présente, au-dessous d'un horizon limono-argileux jaunâtre de dix centimètres d'épaisseur, une couche noire de charbons de bois reposant sur une bande rouge brique d'un centimètre (terre cuite). Puis le sol se poursuit par dix centimètres d'un limon jaunâtre et l'on retrouve une couche de charbons plus émiettés, d'un noir moins vif. Enfin, sous une nouvelle couche de limons se trouve un troisième lit de charbons plus petits enrobés de matière minérale.
On peut conclure de cette observation que le premier feu, des sédiments ont recouvert la surface charbonneuse et que le deuxième feu a été allumé plus tard à la surface du nouveau sol et ainsi de suite.

Plusieurs générations de charbons se trouvent séparées par des apports limoneux.
La couche la plus profonde a 1000 ans.
Ces feux, qui ont duré suffisamment longtemps pour cuire la terre et abandonner de nombreux charbons, ont accompagné les phases d'exploitation de la forêt.
Pour préciser cette hypothèse, des charbons de chacune des couches ont été datés par la méthode au carbone 14. Les résultats donnent pour les plus profonds un âge absolu de 1030 ± 190 ans. Ceci pourrait correspondre aux premiers défrichements importants des XIe et XIIe siècles. On en déduit que le sol de la doline s'est épaissi par apports successifs de trente centimètres en mille ans.
Près de Besançon, des poteries romaines ont été retrouvées dans une doline sous soixante centimètres de sol, ce qui correspond à un remplissage du même ordre.
Les sols des dolines sont lessivés
Les sols profonds qui occupent le fond des effondrements karstiques reçoivent, en plus de l'eau de pluie, les eaux de ruissellement de leur bassin d'alimentation et sont donc soumis à un lessivage intense.

Fond d'une doline. On aperçoit l'exutoire qui communique
avec le réseau karstique
Dans les conditions normales d'un sol de plateau, rappelons que le lessivage se traduit entre autres par un appauvrissement en cations et en argiles dans la partie supérieure du sol et un enrichissement en ces mêmes éléments dans la partie profonde. L'arrivée d'argiles en suspension provoque autour des unités structurales de l'horizon profond un dépôt très fin, très régulier, d'argiles "orientées" qui enrobent les polyèdres de pellicules irisées appelées argilanes. Ces dépôts se distinguent à l'œil nu grâce à l'aspect satiné qu'ils confèrent aux polyèdres mais sont particulièrement spectaculaires au microscope. Sur des lames minces de sol, il est possible de reconnaître les différentes phases de lessivage, leur intensité et leur nature (argiles seules ou argiles et matière organique par exemple).
Qu'en est-il dans les sols de dolines ?
L'analyse indique un lessivage de cations. Le calcium, le magnésium, le potassium sont deux à cinq fois plus abondants dans l'horizon profond que dans l'horizon moyen (l'horizon de surface étant légèrement enrichi par les "remontées biologiques", c'est-à-dire par la bioturbation due à la faune du sol (vers de terre en particulier).
L'observation du profil permet de déceler des argilanes dans l'horizon profond, mais ces dépôts ont un caractère inhabituel. Ils sont irréguliers et parfois ponctués de petites cupules. Au microscope, on en distingue plusieurs générations. Les premières sont cassées et brassées dans le matériau, tandis que les plus récentes semblent encore fonctionnelles, mais présentent des anomalies que nous décrirons plus loin.
L'analyse granulométrique apporte cependant une surprise de taille : il n'y a pas d'enrichissement en argiles en profondeur ! Le profil tout entier présente à cet égard une remarquable homogénéité. Par exemple, dans une doline du Massif du Risol, on a même un horizon profond dont tous les caractères morphologiques sont ceux d'un niveau d'accumulation et qui est le plus pauvre en argiles. Or sous cet horizon on a découvert l'exutoire de la doline en débarrassant un empilement de pierres et de blocs. Ce trou de quelque vingt centimètres de diamètre, en relation avec les conduits profonds du karst, laissait même s'échapper un courant d'air capable d'éteindre la flamme d'un briquet !
Ce cas très démonstratif illustre bien la situation particulière des sols des dolines, disposés sur une sorte de passoire et prêts à s'échapper par les trous.
Les sols s'évacuent par le fond
L'érosion des argilanes
Si l'on regarde attentivement les argilanes déposés dans les horizons profonds, on peut remarquer l'irrégularité du dépôt. Certaines plages montrent des creusements qui ont repris et usé un dépôt antérieur dont il ne reste que des becquets témoins. Ceci explique pourquoi l'horizon dit d'accumulation ne contient pas plus d'argiles que les autres. Il constitue en réalité un horizon de transit dans lequel l'appel au vide dû au soutirage karstique sous-jacent crée l'effet tourbillonnaire qui érode en cupules les faces des polyèdres.
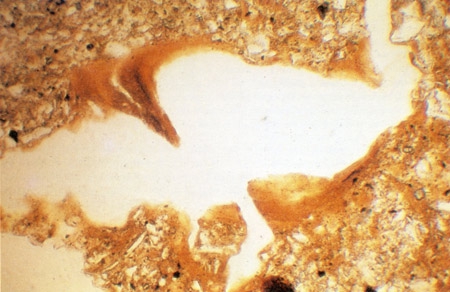
L'érosion a enlevé ce que le lessivage avait apporté. Il ne reste que des becquets témoins
La répartition des charbons et des concrétions
Les charbons de bois sont toujours abondants dans les sols de dolines. À côté de ceux qui proviennent de feux allumés dans les bas-fonds, on trouve également ceux qui ont été apportés par les eaux de ruissellement grâce à leur faible densité. Or ces "traceurs", témoins des apports, ne sont pas seulement concentrés en surface, mais se trouvent disséminés à toutes les profondeurs.
Il en est de même des concrétions ferrugineuses patinées qui constituent la majorité des "sables" des dolines de basse altitude.
C'est donc le profil tout entier qui est concerné par les apports et ces apports s'enfoncent à mesure que l'exutoire avale les horizons profonds.
L'absence de matériau ancien dans les dolines
Un certain nombre de sols de la Forêt de Chailluz ont fait l'objet d'études minéralogiques sur les éléments fins inférieurs à 16 µm. Les recherches ont porté sur des "toposéquences", c'est-à-dire des combinaisons réunissant un sol de plateau et celui d'une doline voisine.
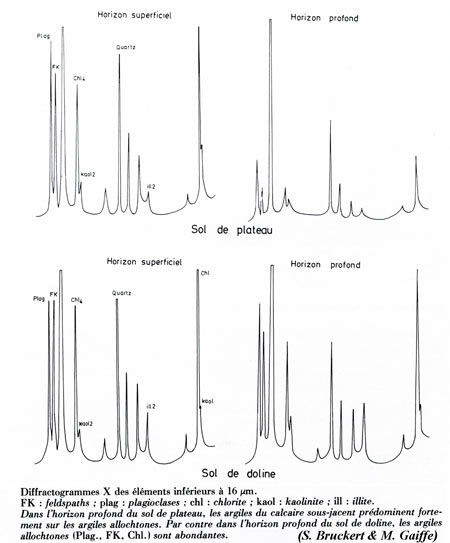
Sur le plateau on met en évidence deux couches bien différenciées : la plus profonde renferme tous les types d'argiles qu'on peut trouver à l'état d'impuretés dans le calcaire du socle et correspond à une argile de décarbonatation. La plus superficielle renferme encore les mêmes minéraux mais aussi — et en quantité notoire — des minéraux (qui n'existent pas dans les calcaires du Jura (feldspaths, plagioclases).
Dans les dolines, ces minéraux allochtones se rencontrent dans toute l'épaisseur du profil et on ne retrouve pas la couche d'argile que le calcaire a pourtant libérée en se dissolvant massivement.
Dans la logique des observations précédentes, ce sont en effet les matériaux du fond du sol — donc les plus anciens — qui sont les premiers avalés par les exutoires karstiques et le sol subit un enfoncement progressif.
La descente des concrétions ferrugineuses
Dans les sols anciens de Basse Franche-Comté, on trouve des concrétions ferrugineuses patinées qui sont héritées de phases d'altérations tertiaires et qui ont été redistribuées. On en dénombre environ deux fois plus dans les sols des dolines que dans ceux des plateaux, ce qui s'explique par une concentration sélective lors du départ des éléments fins.

Dans la partie supérieure des sols de dolines il se produit, lors du dégel printanier, une phase d'engorgement temporaire qui se traduit par une réduction du fer : l'horizon de surface prend une teinte gris-bleu. Au début de l'été, le sol se ressuie et s'aère ; le fer s'oxyde, se concentre en des points privilégiés et forme soit des cylindres rouilles engainant les radicelles, soit des concrétions à structure concentrique autour d'un quelconque noyau.
Lessivés, ces nodules relarguent donc du fer dans les eaux du réseau karstique. Il n'est donc pas étonnant que la nappe phréatique où s'alimentait autrefois la source Marguerite de Thise soit chargée en sels de fer.
Cette partie superficielle du sol affectée par le gel et l'hydromorphie (stagnogley) est la seule zone de fabrication des concrétions. Parmi ces dernières, les gaines racinaires sont des formes transitoires qui se font et se défont sur place, selon les conditions physico-chimiques saisonnières. Les concrétions à structure concentrique par contre sont stables. Si elles ne se forment jamais dans les horizons sous-jacents, elles s'y trouvent transportées par les mouvements d'enfoncement du sol. Au cours de la descente le long du profil, elles subissent une transformation progressive et dans l'horizon profond elles ne conservent souvent qu'un noyau ferro-manganique noirâtre tandis que les couches externes sont décolorées et poreuses. À ce niveau en effet, le léger confinement du sol provoque une remise en solution du fer qui abandonne le cortex de la concrétion.
Contrairement aux argiles qui peuvent transiter à travers le profil grâce aux solutions gravitaires, les concrétions ne peuvent descendre vers le karst qu'accompagnées par l'ensemble du sol. Leur présence jusqu'au fond, alors qu'elles sont fabriquées en surface, illustre parfaitement ce phénomène.
En conclusion, on peut donc affirmer que les sols des plateaux calcaires, malgré leur apparente stabilité, sont en perpétuel mouvement. On peut décomposer leurs déplacements en deux phases. D'abord, comme partout ailleurs, les sols des points hauts ont tendance à être entraînés vers les bas-fonds : ceci se passe d'une façon discrète et continue par la mise en suspension et le transport de certains éléments fins dans les eaux de ruissellement. Lors de la conjonction de facteurs défavorables (déforestation, labour, action des campagnols), suivis d'orages violents, au contraire, le phénomène peut être massif et spectaculaire. Ce ne sont plus les argiles qui se déplacent mais des agrégats intacts et parfois même des masses de sol. Les dolines jouent le rôle de bassins de réception pour tous ces éléments, ce qui explique certains caractères colluviaux de leurs profils : épaisseur, homogénéité, porosité, présence d'éléments hérités (charbons, concrétions).
Mais ce qui fait l'originalité de ces zones karstiques, c'est que les bas-fonds ne sont pas que des bassins de réception. Les dolines étant en communication avec les réseaux souterrains, il se crée un appel au vide qui déstabilise perpétuellement le remplissage et tend à l'aspirer vers les profondeurs. Bruckert et Gaiffe ont pu montrer que cette usure par le fond a déjà avalé, à l'étage collinéen, tous les vestiges des sols anciens et que ce phénomène se poursuit.
Entre ces deux mouvements, il peut ne pas y avoir une équivalence parfaite. Il est probable qu'un grand nombre d'anciennes dolines à l'exutoire étroit ont reçu plus de matériaux qu'elles n'en ont exportés. De ce fait, elles se trouvent comblées et n'apparaissent plus dans le paysage. On reconnaît leur existence lorsque des travaux de terrassement mettent à jour ces énormes poches de terre.
Dans certains cas plus rares, l'exportation vers la profondeur est supérieure à l'alimentation. Il subsiste alors un entonnoir, au fond duquel des cailloux lavés marquent remplacement de la perte. De tels exemples montrent la puissance potentielle de ce type original d'exportation des sols.
Actuellement l'érosion reste limitée dans ces secteurs de plateaux grâce à la présence de la prairie qui assure une couverture à peu près complète et une protection efficace du sol. Le phénomène se déclenche cependant chaque fois que la terre est mise à nu : au moment des labours, avec la pullulation des petits mammifères qui bouleversent la surface, avec les exploitations intensives en forêt et les coupes à blanc.
Sources :
Bruckert S. & Gaiffe M. (1985). — Les sols de Franche-Comté CUER Université de Franche-Comté 142 p.
Rolin P. (2016). — Livre guide d'une excursion géologique en forêt de Chailluz.
22:12 Publié dans Botanique, Géologie-hydrogéologie-Climatologie | Tags : dolines, thise, forêt de chailluz | Lien permanent | Commentaires (0) | Facebook | | |
Les sols de la forêt de Chailluz
Les sols de la forêt de Chailluz
(dernière mise à jour : avril 2015)
Généralités
Au cours des temps historiques, la forêt comtoise était habitée de bûcherons, de charbonniers, de forgerons et de leveurs d’écorces, sans compter les chasseurs, braconniers et simples usagers. Peuplée de jour comme de nuit, théâtre d’une activité intense, elle était largement exploitée.
Si la Franche-Comté est aujourd’hui couverte à plus de 40 % de résineux et de feuillus en tout genre, on sait de manière certaine que ce taux était largement inférieur entre la fin du Moyen-Âge et le XVIIIe siècle. Ici comme ailleurs, la déforestation devient massive à partir du XVe siècle pour répondre à la fois aux besoins en énergie et en matériau de construction.
« Il ne faut pas oublier que la région est riche d’une tradition industrielle pluriséculaire, rappelle Paul Delsalle, historien à l’université de Franche-Comté[1], et que dès le Moyen-Âge, les usines comptent parfois jusqu’à deux ou trois cents ouvriers ! » Les salines disséminées sur tout le territoire en sont des exemples. Avec près de mille ouvriers au XVIIe siècle, celle de Salins-les-Bains (39) est sans conteste la plus importante. Trois mille cinq cents hommes s’emploient à la fournir en bois régulièrement !
L’exploitation de la forêt est soumise à des contraintes et des règles précises, cela des siècles avant le rattachement du Comté au royaume de France en 1678 et l’adoption des règles édictées par Colbert en matière de gestion forestière.
Certaines essences étaient réservées à l’industrie, et le droit des habitants se limitait en général au « mort bois » comprenant tilleul, noisetier et charme. Il était interdit de se servir en fruitiers, qui, outre les pommiers, poiriers, pruniers et cerisiers qui abondaient en forêt, comptaient aussi le chêne et le hêtre. Les dossiers de justice fourmillent de condamnations comme celle, au XVIe siècle, de ce Bisontin de retour de Chailluz, arrêté porte de Battant avec un chariot chargé de branches de cerisier. « Mais est-ce que nous ne surestimons pas la présence de ces variétés du fait qu’elles sont régulièrement citées dans les archives ? », se demande Paul Delsalle, qui voit d’un bon œil l’apport d’autres disciplines pour compléter les sources documentaires.
Propriété de la ville de Besançon depuis des temps immémoriaux, la forêt de Chailluz s'étend sur 1673 ha formant un massif compact de plus de 4000 ha avec les forêts adjacentes, toutes communales. Parmi les grandes villes françaises, Besançon est la seule à posséder en toute propriété une forêt d'une grande étendue très proche de l'agglomération.
La forêt de Chailluz constitue l'essentiel du patrimoine forestier de la cité de Besançon qui offre une étendue exceptionnelle de 2073 ha. Une légende attribue à une "Dame de Chailluz" la donation de cette forêt à la ville, sous réserve que les produits en seraient distribués aux pauvres.
La réalité historique est tout autre. Depuis la délimitation du territoire de Besançon en 1442, la propriété de la forêt avait été reconnue à la Ville pour être ensuite contestée pendant trois siècles par les Ducs de Bourgogne, jusqu'à ce qu'un arrêté du Parlement pris en 1705 mette définitivement fin à cette querelle alimentée par les ressortissants des paroisses voisines. La difficulté à établir de façon précise des limites de propriété à l’intérieur des forêts n’est pas sans générer des tensions qui parfois tournent au pugilat. La forêt de Chailluz n’échappe pas à la règle et les Bisontins du XVIe siècle sont à couteaux tirés avec les habitants de Tallenay, Bonnay, Vieilley, Braillans, Chalezeule ou encore de Chatillon-le-Duc dans la défense de leurs lopins communaux. Une réalité d’autant plus criante que l’exploitation de la forêt est capitale à cette époque. Les habitants de Tallenay plantent même du Gamay sur les coteaux sylvestres en 1609. Mais la vigne s’avère difficile à entretenir, le vin de piètre qualité, et devant une production qu’il juge excessive, le Parlement de Dole ordonne l’arrachage des ceps. Le vin de Chailluz ne sera plus conservé que dans des pages d’archives…
Céline Bouvresse[2] est enseignante en histoire et travaille régulièrement sur les forêts comtoises au travers de travaux de recherche universitaires. « Au XVIe siècle, les limites étaient fixées grâce à des points de repères naturels comme la crête d’une colline, ou d’autres plus discutables car potentiellement changeants : le tracé d’un chemin, la pose d’une borne en pierre ou la gravure d’un emblème sur un arbre. Les descriptions n’étaient qu’orales et on apprenait aux enfants à reconnaître les lieux. Il n’est pas rare que les dossiers de justice s’appuient sur les témoignages des anciens du village faisant appel à leurs souvenirs d’enfance pour servir de preuve. » Il faudra attendre le début du XVIIIe siècle pour que les premiers plans apparaissent et limitent les conflits en même temps que les propriétés.
Traitée pendant des siècles en "taillis sous futaie" en vue de la production de bois de chauffage (taillis) et accessoirement de bois de construction (futaies = troncs des gros arbres provenant des tiges réservées au moment des coupes périodiques du taillis), cette forêt donnait lieu à l'exploitation annuelle d'une coupe de 42 ha et d'un coupon de 14 ha. Les milliers de stères de bois étaient façonnés par une Régie municipale employant une grand nombres de bûcherons et de voituriers. Les habitations construites aux Grandes Baraques étaient autrefois plus nombreuses et destinées au logement des bûcherons de la Ville. Le régisseur revendait le bois au prix coûtant pour tenter de satisfaire les énormes besoins de la Ville en bois de feu. Cette régie municipale existe toujours, mais elle joue un rôle plus limité, la plupart des coupes étant vendues sur pied à des exploitants ou des industriels du bois.
Aurore Dupin est une jeune chercheuse spécialiste en anthracologie, l’étude des charbons de bois. Doctorante au laboratoire Chrono-environnement et rattachée à la MSHE où elle prépare une thèse en archéologie, la forêt de Chailluz s’avère pour elle un excellent terrain d’investigation depuis que la technologie LIDAR (télédétection par laser aéroporté) a révélé les traces d’un millier de charbonnières, dédiées précisément à la fabrication du charbon, dont les résidus permettront d’identifier les essences d’origine.
« De nombreuses informations nous proviennent de la forêt de Chaux, où l’on perpétue encore la tradition du travail des charbonniers, explique Aurore Dupin. Pour la forêt de Chailluz, il n’existe plus de mémoire, peu de documents et pas de vestiges d’habitations qui toutes étaient construites en matériaux périssables. » Les méthodes scientifiques aident à pallier ce déficit. La susceptibilité magnétique confirme dans un premier temps les relevés du LIDAR. Elle certifie que l’argile du sol a subi des températures extrêmes. « Lorsque l’on chauffe fortement de l’argile, les minéraux qui la composent s’organisent d’une manière particulière, guidés par le champ magnétique terrestre. » La datation au carbone 14 atteste ensuite l’existence de la majeure partie des vestiges entre le XVIIe et le XIXe siècles. À partir d’infimes résidus, le microscope optique à réflexion est capable de déterminer l’essence du bois grâce à des caractéristiques anatomiques que le charbon présente sur trois faces. Paul Delsalle aura peut-être dans les mois qui viennent des réponses quant à la présence des fruitiers en forêt sur laquelle il s’interroge…

Charbonnière en forêt de Chaux - Le bois était empilé en meule selon une géométrie étudiée, puis recouvert de terre de façon à garantir une combustion « à l’étouffée ». Le feu était surveillé jour et nuit. Une charbonnière atteignait en moyenne six mètres de diamètre et deux mètres de hauteur, et représentait cinq à six stères de bois. © En direct n° 254 septembre-octobre 2014
[1] Paul Delsalle - Aurore Dupin Laboratoire Chrono-environnement Université de Franche-Comté Tél. (0033/0) 3 81 66 58 74
[2] Céline Bouvresse Tél. (0033/0) 6 83 24 90 78
Aperçu sur la flore et la faune de la forêt de Chailluz
Poussant sur un sol souvent ingrat, la forêt de Chailluz est relativement pauvre, mais elle a été progressivement enrichie grâce à la vigilance et aux efforts conjugués de la Municipalité et des forestiers.
La zone nord-est a été longtemps occupée par des cultures comme le prouvent les vestiges de murets et d'amas d'épierrement. La forêt de Chailluz a donc progressé en superficie comme une hêtraie pratiquement pure. Pendant des siècles, les traitements qui lui ont été appliqués ont favorisé le Charme et le Chêne. De sorte que le Chêne occupe une place importante même dans certaines zones qui ne lui sont pas particulièrement favorables. Il faut dire que cette essence a été longtemps favorisée au détriment du Hêtre grâce à une sélection pratiquée lors des coupes, et que pendant très longtemps on a planté quantités de Chênes. Au début du XXe siècle, on plantait chaque année 10 000 Chênes !
Autres espèces arborescentes feuillues rencontrées : du Frêne, des Érables, du Tremble, du Merisier, du Bouleau, de l'Alisier. L'Orme a pratiquement disparu à cause de la graphidiose. Les résineux ont tous été introduits : de l'Épicéa, en grand nombre ainsi que des Sapins et quelques Pins.
Les flores arbustive et herbacée caractéristiques des collines calcaires sont bien représentées. Et dans certaines parcelles (parcelles 8 à 16), la présence d'argiles à chailles favorise quelques espèces calcifuges comme le Châtaignier et la Fougère Aigle.
La macrofaune comporte des sangliers, des chevreuils, des renards, des blaireaux et une vingtaine d'espèces d'oiseaux.
Qu'est-ce que la pédologie ?
Les sols constituent l'élément essentiel des biotopes propres aux écosystèmes continentaux. Leur ensemble, dénommé pédosphère, résulte de l'interaction de deux compartiments biosphériques : l'atmosphère et les couches superficielles de la lithosphère. La pédogenèse représente la formation des sols et l'étude des sols est une science qui s'appelle la pédologie. Les ressources de notre environnement n'étant pas inépuisables, on se doit de les utiliser au mieux. Cet impératif passe obligatoirement par une gestion rationnelle des forêts et des terres, ce qui nécessite une connaissance approfondie des qualités actuelles des sols, des mécanismes capables de les dégrader et de les détruire, et des moyens de les conserver et de les améliorer. Les différentes couches de matériaux homogènes qui constituent le sol sont appelées horizons en pédologie.
La formation des sols représente un processus complexe consistant en la transformation des roches situées à la surface de la croûte terrestre (roches mères) par l'effet conjugué des facteurs climatiques et des êtres vivants. Il est en réalité impossible de comprendre la genèse des sols si l'on ne prend pas en considération le rôle des organismes : bactéries, champignons et autres cryptogames, plantes vertes, pédofaune.
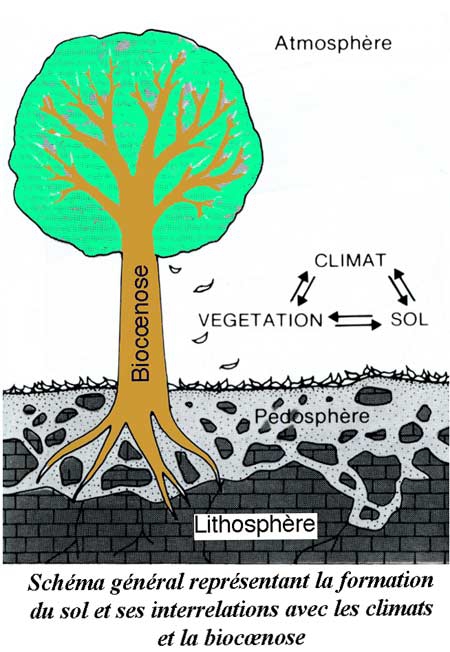
L'altération des roches mères commence par un phénomène de désagrégation physique. Celui-ci est provoqué par faction des facteurs climatiques : variations nycthémérales de température, érosion hydrique à laquelle s'ajoute ultérieurement la fracturation du substratum rocheux par les racines des végétaux. Un processus de décomposition chimique qui fait suite, induit par le lessivage qu'effectuent les eaux d'infiltration chargées de substances dissoutes (CO2 par exemple) qui solubilisent la roche et aussi par les sécrétions corrosives de divers végétaux pionniers. L'ensemble de ces processus fragmente la roche mère et la transforme chimiquement en la dissociant en ses composés initiaux.
En définitive, les sols résultent de l'action extrêmement intriquée et complexe des facteurs abiotiques et biotiques qui conduit à l'élaboration d'un mélange intime de matières minérales et organiques provenant de la décomposition des êtres vivants après leur mort et de leurs excréta (litière, racines mortes, cadavres d'animaux, fèces, etc.)
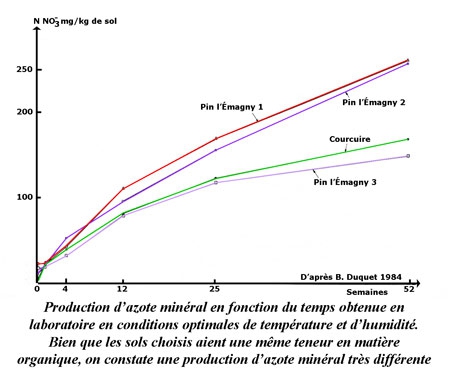
Le sol met des milliers d'années pour se former. C'est un bien précieux qui peut disparaître avec une facilité et une rapidité déconcertantes. Les exemples ne manquent pas. Dans le monde, des surfaces considérables de terres fertiles sont irréversiblement perdues chaque année par érosion, sur-exploitation ou à la suite d'aménagements inappropriés.
Caractères physiques des sols
Les principaux facteurs édaphiques sont constitués par la texture et la structure des sols, leur hygrométrie, leur pH et leur teneur en éléments minéraux.
Tous les sols comportent deux fractions distinctes l'une minérale, l'autre organique, intimement mélangées en un complexe organo-minéral. La texture dépend de la nature des fragments de roche mère ou de minéraux provenant de sa décomposition que renferme la fraction minérale. L'analyse granulométrique permet de distinguer dans cette dernière des éléments grossiers (cailloux et graviers) ainsi que des éléments fins (sables, limons et argiles).
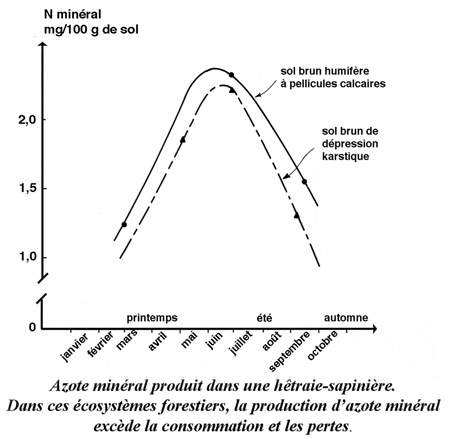
Les cailloux sont de taille supérieure à 20 mm, les graviers mesurent entre 2 et 20 mm de dimension maximale, les sables de 2 mm à 20 µm de diamètre, les limons de 20 µm à 2 µm, les argiles moins de 2 µm.
La proportion relative des éléments fins constituant la fraction minérale permet de classer selon leur texture les divers types de sols. Elle présente une grande importance agronomique et de façon plus générale pour l'ensemble des écosystèmes terrestres car c'est d'elle que dépend pour une grande part la circulation de l'eau dans les sols.
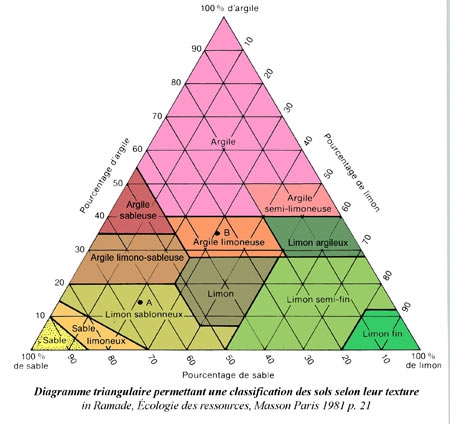
Structure des sols
L'architecture des sols dépend de l'état des particules qui les constituent. Lorsque les particules les plus fines, de nature colloïdale, sont floculées, elles forment des agrégats en cimentant les éléments de plus grande taille entre lesquels existent des lacunes.
Si, à l'opposé, elles sont dispersées, les éléments du sol vont rester indépendants et ne délimiteront pas de système lacunaire bien défini. Les sols du premier type sont dits en agrégats, ceux du second type, particulaires.
La porosité constitue un autre paramètre édaphique important qui combine les critères propres à la texture et à la structure du sol considéré. La porosité peut se définir comme la proportion du volume des lacunes par rapport au volume total.
De cette dernière dépend la circulation de l'eau et des gaz dans les sols dont le rôle est essentiel aussi bien pour assurer le développement des plantes supérieures que celui de la microflore et de la faune édaphique.
La porosité décroît lorsque l'on passe de structures en agrégats très lacunaires vers des structures de plus en plus particulaires. Lorsque les sols particulaires sont dépourvus de sable, ils peuvent devenir asphyxiants car ni l'eau ni les gaz ne peuvent y circuler normalement.
Formation des sols ou pédogenèse
La pédogenèse résulte de l'action des facteurs écologiques abiotiques et biotiques sur les couches supérieures de la lithosphère.
La formation des sols commence par la fragmentation de la roche mère suivie d'une seconde étape marquée par la corrosion des minéraux présents. Celle-ci résulte de processus complexes : oxydation, réductions, hydratation, hydrolyse, etc. Le lessivage provoqué par les pluies et des facteurs topographiques (sols en pente ou sols plus ou moins bien drainés) va mettre en solution les produits de ces réactions chimiques. Le processus est favorisé par l'action des végétaux pionniers. Des cryptogames, tels les lichens exercent par leurs sécrétions une action corrosive intense sur les minéraux constitutifs des roches. De plus, les racines des plantes pionnières, outre qu'elles fissurent le substratum rocheux, accélèrent la dissolution des minéraux par leurs exsudats, conjointement au CO2 dissous dans l'eau d'inhibition.
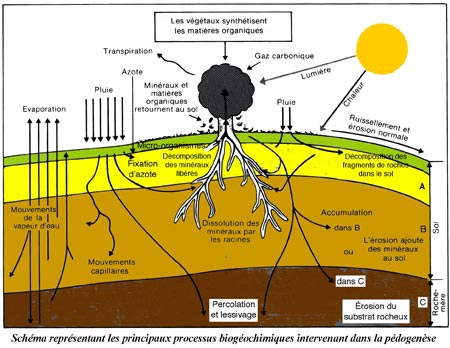
Si l'érosion des sols est apparemment réduite en Franche-Comté, c'est d'abord le résultat des aménagements judicieux menés pendant des générations, à quelques. La forêt primitive a généralement fait place à un bon équilibre agro-sylvo-pastoral. Les cas d'aménagements mal appropriés proviennent la plupart du temps d'une méconnaissance des qualités des sols et de leur environnement.
La monoculture d'une espèce végétale, que ce soit en milieu agricole ou forestier, est toujours néfaste à plus ou moins longue échéance. La monoculture d'une essence forestière diminue obligatoirement l'activité des microorganismes. Ceux-ci en effet se trouvent fortement stimulés quand les aliments proviennent de sources diverses, mais n'apprécient guère le "plat unique". De ce fait, le cycle biologique se ralentit et les éléments nutritifs, les cations notamment, se retrouvent bloqués dans les débris organiques qui se décomposent plus lentement, d'où une acidification du sol.
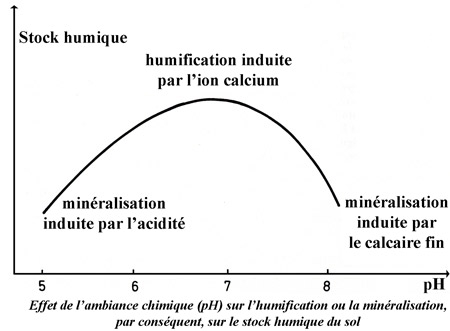
Les résineux sont souvent accusés de dégrader les sols : comme nous le verrons plus loin, les feuillus comme le hêtre et le chêne sont tout aussi dégradants s'ils sont cultivés en peuplement pur. La dégradation des terres est en fin de compte liée bien davantage aux propriétés intrinsèques des sols et aux actions d'aménagements qu'aux résineux eux-mêmes.
Formation de l'humus
Elle s'effectue par addition de matière organique aux constituants minéraux des sols et représente la troisième phase de la pédogenèse dont la responsabilité est dévolue en premier lieu aux êtres vivants. Ce processus est donc contrôlé essentiellement par les facteurs biotiques.
La matière organique incorporée aux sols provient essentiellement de la litière, laquelle est constituée de feuilles mortes, de brindilles et d'autres fragments végétaux. Les branches et les troncs morts et, dans une bien moindre mesure, les excréments des herbivores contribuent à cet apport de matière organique.
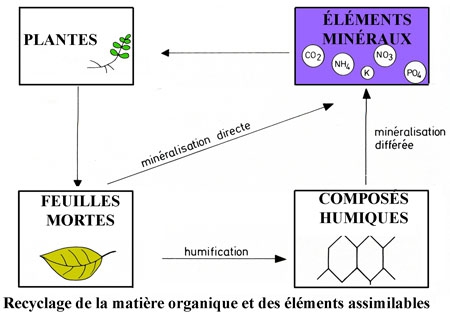
Par le jeu de la photosynthèse, les végétaux apportent beaucoup plus de matières aux sols qu'ils n'en prélèvent. En tout état de cause, les éléments minéraux absorbés par les racines sont restitués lors de la mort des producteurs primaires. Certains micro-organismes, telles les bactéries fixatrices d'azote, incorporent des nitrates aux sols à partir de l'azote atmosphérique. En conséquence, l'activité des plantes supérieures et de certaines bactéries édaphiques apporte au sol plus de matière qu'elle ne leur en enlève. En outre, cette matière est généralement amenée sous forme de dérivés organiques complexes dont certains ne se décomposent que très lentement.
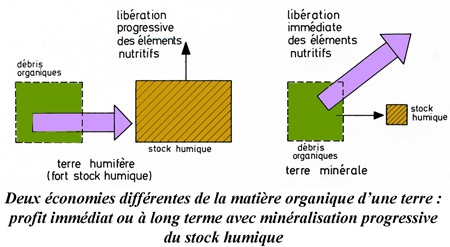
Les facteurs biotiques vont intervenir de façon déterminante dans la dégradation des matières organiques mortes.
Une première phase est sous la dépendance de nombreuses espèces animales qui vivent à la surface ou à l'intérieur des sols. Celles-ci interviennent directement et (ou) indirectement pour fragmenter la matière organique et pour l'introduire dans les couches profondes.
Certaines espèces, comme les rongeurs terricoles, amènent des résidus provenant de l'accumulation des détritus végétaux à l'intérieur des sols par suite de leur activité de fouissage.
À l'opposé, les invertébrés saprophages jouent un rôle pionnier dans la formation de l'humus en fragmentant la matière végétale morte : litière, branchages, fragments de bois dont ils se nourrissent dans le cas des termites. Il en est de même des coprophages, qui s'alimentent des excréments de vertébrés, en particulier d'ongulés, dont le rôle est essentiel dans les écosystèmes prairiaux (savanes, steppes, etc.).
Par son activité, la faune du sol (pédofaune) disperse dans les couches profondes la matière organique morte présente en surface et ramène dans les couches superficielles leurs excréments contenant les produits de digestion de la litière.
La pédofaune est constituée par une grande variété de groupes taxonomiques d'invertébrés. Parmi ces derniers, on dénombre des arthropodes, des annélides oligochètes, des mollusques, des crustacés isopodes (cloportes) et divers autres phyla ; rotifères, nématodes, protozoaires, etc.

Principaux types d'organismes saprophages (invertébrés) et décomposeurs (bactéries, champignons) constituant les peuplements de la litière et des sols.
A, B, C, collemboles gen. Tomocerus, Isotoma, Folsomia ; D, larve de bibionide (diptère) : E, diploure ; F, lombric (annélide oligochète) : G, Lithobius (chilopode) : H, Glomeris ; I, iule (diplopodes) ; J, nématode ; K, L. acariens Oribatides gen. Belba et Oribotridia ; M, N, R, champignons phycomycètes (Rhizopus), ascomycètes (Aspergillus) et Pénicillium ; O, streptomycètes ; P et Q bactéries : Cytophaga (cellulolytique aérobie ; Clostridium (fixateur d'azote, anaérobie). D'après Duvigneaud, La synthèse écologique, 2e édition, 1980, mais modifié. Doin, Paris.
Par leur biomasse, les oligochètes (lombrics ou vers de terre) et, par leur nombre, les arthropodes constituent les deux groupes dominants. Ces derniers sont représentés par des acariens oribatides, des myriapodes (diplopodes et chilopodes), des aptérygotes (surtout collemboles), des insectes (surtout des larves de diptères et de coléoptères).
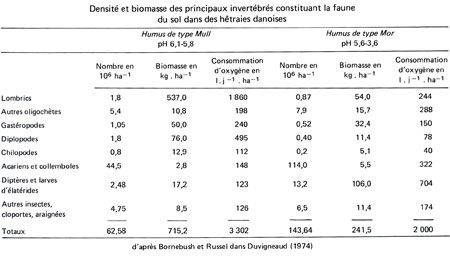
En définitive, la biomasse constituée par la pédofaune est aussi considérable que le nombre d'individus qu'elle comporte à l'hectare. Il est par exemple banal de dénombrer dans une forêt caducifoliée tempérée plusieurs dizaines de millions d'arthropodes par hectare. De même, la biomasse de vers de terre, en moyenne de l'ordre de 500 kg/ha, peut atteindre plus de 1 000 kg/ha dans des sols forestiers très riches en matière organique et même dépasser 2800 kg/ha dans certaines prairies tempérées ! Comme le fait remarquer fort à propos Duvigneaud, en Europe occidentale pourtant surpeuplée, la biomasse des lombrics dépasse celle des hommes !
Types d'humus
La formation de l'humus, conditionnée en premier lieu par l'activité biologique des sols, dépend aussi de facteurs physicochimiques — dont certains ont d'ailleurs une forte interdépendance avec les facteurs biotiques — aération, teneur en eau édaphique (drainage), pH, nature du substratum rocheux, etc. Ainsi le type d'humus constitue-t-il une caractéristique essentielle car il intègre l'influence de l'ensemble des facteurs écologiques propres au biotope considéré.
On distingue quatre types d'humus en fonction de la rapidité de dégradation de la litière et de la décomposition des matières organiquesqui en dérivent : les Mull, les Moder, les Mor et les tourbes.
La formation des Mull prend lieu dans les sols riches et profonds, frais, bien drainés donc bien aérés. Ici, la décomposition de la litière est très rapide par suite de la grande abondance de la pédofaune, en particulier des vers de terre dont l'activité assure en outre une excellente dispersion de l'humus dans l'horizon[1] supérieur du sol. Les lombrics ingèrent en effet un mélange de fragment de litière et de terre. On estime que pour une densité de 50000 lombrics par hectare, ces annélides "consomment" quelque 2,5 tonnes de feuilles mortes pendant la belle saison mélangées à 25 tonnes de terre. On évalue en moyenne à 42 tonnes par an la masse de déjections produites par ces invertébrés[2]. En une trentaine d'années, les premiers décimètres du sol sont donc entièrement "digérés" par les lombrics. Si l'on ajoute à ces considérations le fait que les sécrétions muqueuses des vers de terre sont indispensables à l'activité de nombreuses bactéries du sol, en particulier de celles qui fixent l'azote, on imaginera bien l'importance écologique capitale de ces invertébrés dans les écosystèmes terrestres tempérés).
Par suite de l'intense activité des lombrics et autres animaux saprophages, l'horizon A1des Mull forestiers est caractérisé par une excellente homogénéisation des matières organiques en décomposition conférant à cet horizon une teinte plus foncée.
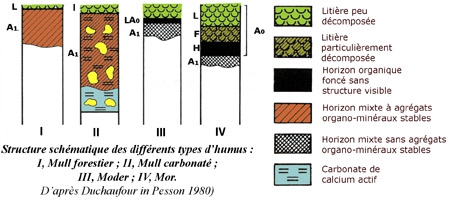
Dans les écosystèmes où prédominent les végétaux herbacés croissant sur sol calcaire, il se forme un Mull carbonaté. Ici la décomposition de la litière est un peu moins rapide que dans les Mull forestiers, d'où formation d'un horizon A1, épais et foncé.
Les Moder correspondent à des humus très répandus sur les sols pauvres. Comme dans le cas des Mull, il s'agit d'un humus dit coprogène car constitué essentiellement d'une poudre brune provenant des déjections globuleuses de divers invertébrés coprophages, dont le rôle consiste à déchiqueter la litière, à la fragmenter ainsi que les autres débris végétaux dont ces animaux se nourrissent.
Ici, le rôle des arthropodes est prépondérant, en particulier sur sols squelettiques où les myriapodes dominent au détriment des acariens et collemboles. A l'opposé, les lombrics sont moins bien représentés que dans les Mull. En conséquence, la litière non encore décomposée et celle en voie de dégradation sont beaucoup plus abondantes dans les sols à Moder, constituant un horizon A1 bien différencié.
Peu à peu ramollie par l'eau d'imbibition, la litière des Moder est envahie par une multitude de métazoaires inférieurs (rotifères, tardigrades, nématodes), de protozoaires (rhizopodes et flagellés) sans omettre une myriade d'algues microscopiques et de bactéries qui prolifèrent dans le compost ainsi formé.
Ce compost est en outre le siège d'un intense développement fungique, un abondant feutrage mycélien envahissant la matière organique en voie d'humification.
Les Mor constituent un autre type d'humus dit mycogène car sa production est essentiellement assurée par des champignons saprophytes. Les Mor se rencontrent souvent sur des sols très pauvres, installés sur une roche mère granitique, dans des landes à bruyères ou sous toute autre formation végétale acidifiante (forêt de conifères par exemple), et lorsque les conditions climatiques sont défavorables (froid humide).
Les Mor apparaissent en définitive sur des sots marqués par une faible activité biologique. L'importante accumulation de la litière se traduit par l'existence d'un horizon A0 très épais. Elle résulte de la rareté des animaux saprophages, lombrics et myriapodes en particulier. À l'opposé s'observe une grande abondance de champignons saprophytes qui peuvent représenter en poids jusqu'à 15 % de l'horizon A0 !
À la base de cet horizon s'accumule une couche organique de couleur brun-noir, fibreuse, dénommée "humus brut", superposée directement au sol minéral car l'horizon A1, est peu développé voire inexistant.
Les conditions d'acidité des Mor défavorisent l'activité bactérienne. De la sorte, la matière organique est mal décomposée et l'humification n'est pas menée à terme. Il se forme surtout des acides fulviques, composés phénoliques partiellement solubles qui sont produits par l'altération des membranes cellulaires. Ces acides, entraînés par les eaux d'infiltration, altèrent les colloïdes des argiles et interdisent de ce fait la formation des agrégats propres au complexe argilo-humique.
Pour mémoire, car il n'y en a pas en forêt de Chailluz, mais dans le Haut-Jura, les tourbes sont constituées par l'accumulation d'une grande quantité de matériaux organiques incomplètement décomposés, caractérisés par un taux d'humification généralement assez faible. Les tourbes se forment dans des milieux saturés en eau de façon quasi permanente — les tourbières — où apparaissent des conditions d'anaérobiose défavorables à toute activité biologique. Par suite de la quasi-absence de la faune et de la grande pauvreté de la flore bactérienne et cryptogamique, la matière organique se transforme très lentement et s'accumule souvent sur plusieurs mètres.
On distingue deux types de tourbes, les tourbes eutrophes qui s'édifient dans les tourbières de fond de vallée, là où la nappe phréatique affleure en permanence et les tourbes acides, quiseforment dans les tourbières de montagne, aux dépens des eaux de pluie qui s'accumulent dans des dépressions, sur sols acides. Dans un cas comme dans l'autre, seuls les horizons superficiels sont partiellement humifiés, lors des périodes d'assèchement de la surface qui en permettent l'aération.
Processus biochimiques de la formation de l'humus
Ces processus caractérisent la transformation de la matière organique morte par les agents microbiologiques, actinomycètes et bactéries, après qu'elle a transité dans le tube digestif des animaux saprophages. Les composés solubles ou facilement solubilisables qui préexistent dans la litière — dénommés composés hérités — vont migrer rapidement dans les sols, de même que les composés solubles néoformés, provenant en particulier de l'hydrolyse de la cellulose. Après séparation dans l'espace des composés insolubles et solubles, ces derniers vont se mélanger aux éléments minéraux, étape qui représente un des facteurs essentiels de l'humification.
Dans les Mull et les Moder, l'action des micro-organismes sur les matières insolubles est différente selon qu'il s'agit de cellulose ou de lignine. En milieu basique, la cellulose est dégradée plus rapidement que la lignine alors que leur vitesse de dégradation est comparable dans les Mull acides et dans les Moder.
Dans les Mull neutres ou basiques, la lignine est transformée en humus brun, insoluble. Il s'y constitue en outre des associations stables lignine-protéines. Dans ces mêmes milieux, l'action des bactéries conduit à des formations de composés cellulose-protéine, qui constituent les acides humiques gris, plus stables et résistants que les précédents. D'autre part, la biodégradation de la lignine, mais aussi la néoformation bactérienne de polysaccharides, conduisent à la production d'un ensemble de composés insolubles, de fort poids moléculaire, désignés sous le terme général d'humine. Celleci arrive à représenter en poids plus de 50 % de diverses fractions constituant l'humus.
Dans les Mull, et aussi dans une moindre mesure dans les Moder, acides humiques et humine se lient aux argiles colloïdales. Il s'édifie de la sorte, grâceà ces "ciments" humiques, des structures composites, dénommées sous le terme général de complexe absorbant argilo-humique, auxquelles sont également associés divers autres composés minéraux : calcaire actif, allophanes, et cations lourds.
On constate que l'activité bactérienne des sols diminue au fur et à mesure que l'humus s'accumule. De la sorte, une certaine quantité des sels minéraux nutritifs (nitrates, phosphates, sels de potassium, etc.) libérés par dissolution et (ou) dégradation de la matière organique deviennent disponibles pour les racines des phanérogames et autres végétaux autotrophes.
Enfin, l'humus est peu à peu minéralisé par l'action des microorganismes de sorte que la teneur des sols en matières humiques correspond à un équilibre dynamique entre biosynthèse et dégradation des divers constituants de ces dernières.
L'écologie des sols de la forêt de Chailluz
La région a été épargnée par les glaciers et possède par conséquent une histoire pédologique longue, datant souvent de l'ère tertiaire. Les roches-mères, au cours des millénaires, se sont altérées de façon continue en fournissant une couche épaisse de matériaux pédologiques. Les roches mères étant diverses, leurs altérites sont également très variées. D'autre part l'altération n'intervient jamais seule. L'érosion s'exerce aussi, avec ses périodes modérées et ses paroxysmes. Certains horizons poreux peuvent ainsi migrer, progressivement ou en masse, vers d'autres positions où parfois ils recouvrent des sols déjà constitués.
Troncatures, transports, recouvrements, affectent inégalement les sols, dont l'histoire devient ainsi non plus une évolution continue mais une succession d'épisodes apportant chacun leur marque. De tels sols formés en plusieurs étapes sont dits polygéniques.
Un climat lessivant à peu près constant depuis le tertiaire
Au cours de la pédogenèse, le climat joue un rôle considérable. On sait qu'à l'ère tertiaire une longue période d'altération a démantelé des masses énormes de calcaire. Cette décarbonatation intense n'a pu s'exercer que grâce à un fort drainage climatique, ce qui suppose des conditions d'humidité et de fraîcheur proches des conditions actuelles.
Mais on a pu mettre en évidence d'importantes fluctuations climatiques. Certaines périodes qui ont permis le développement d'une flore de palmiers et d'espèces méditerranéennes ont été chaudes et sèches. Les minéraux ont subi une altération différente et certains sols ont pris une teinte vive due aux oxydes de fer déshydratés (rubéfaction).
Quant aux périodes froides du quaternaire, elles n'ont pas pu rester sans influence sur les sols situés aux marges des calottes de glace. Les alternances de gel et de dégel ont provoqué un brassage par cryoturbation dont certaines traces sont encore bien visibles : nodules d'argiles arrondis isolés dans des limons, fentes en coin remplies d'éléments blanchis (glosses).
Enfin le climat actuel est encore très favorable à l'entraînement par lessivage puisque les précipitations remportent sur 1'évapotranspiration potentielle, sauf pendant les mois d'été qui accusent un léger déficit hydrique. L'existence de cette relative sécheresse estivale aura, dans les secteurs les mieux exposés, certaines incidences sur l'évolution des sols (rubéfaction actuelle). On remarque aussi que dans cette région de faible altitude, les températures moyennes mensuelles ne descendent pas en dessous de zéro degré.
De nombreux faciès calcaires, à l'origine d'une grande variété d'altérites
o Les argiles de décarbonatation
Les roches mères les plus répandues restent à cette altitude les calcaires jurassiques à faciès durs et relativement purs. Leur dissolution laisse de 1 à 10% d'éléments insolubles, surtout de l'argile et quelques grains de quartz. Ces argiles sont un constituant important des sols. Elles donnent les terra fusca (jaunes) et les terra rossa (rouges) qui emplissent les fissures et tapissent les bancs calcaires.
Un cas particulier est constitué par les calcaires de l'Argovien. Dans la partie Ouest du Jura, ce sont des bancs très riches en impuretés argilo-limoneuses et en rognons siliceux, de quelques centimètres de diamètre, constituant les chailles. La dissolution du carbonate laisse donc un abondant résidu connu sous le nom d'argiles à chailles, bien que les argiles proprement dites y soient beaucoup moins abondantes que la partie limoneuse formée par une poudre de silice.
o Les limons et sables de décarbonatation
Certains faciès argoviens sont des calcaires gréseux formés de grains de silice cimentés par du calcaire. La disparition du ciment laisse alors un résidu limono-sableux fin, très filtrant, formé de silice amorphe (calcédonite) très acide.
o Les limons éoliens ou ruisselés de compositions différentes[3].
Les recouvrements de limons occupent des surfaces importantes sur les premiers plateaux jurassiens et dans les plaines alluviales. Leur origine n'est pas partout élucidée mais on peut cependant reconnaître deux catégories :
— les limons éoliens, identifiés par la présence en quantité importante d'éléments minéraux qui n'existent pas dans les calcaires sur lesquels ils reposent. Ils correspondraient à des nuages de poussières déposés par des vents venus du Sud-Est qui auraient arraché des matériaux fins aux moraines alpines, après les glaciations. De tels transferts de poussières s'observent encore de nos jours à une échelle plus réduite : plusieurs fois par an on peut remarquer sur les voitures stationnées en plein air, une couche jaunâtre que les spécialistes attribuent à des nuages de particules d'origine lointaine.
— les limons ruisselés qui peuvent provenir du remaniement des premiers par les eaux de ruissellement, ou bien être déposés lors des crues importantes dans le lit majeur des rivières.
Les études au microscope électronique à balayage permettent de déceler sur les grains soit des cupules résultant des chocs entre grains dans le cas de l'éolien, soit des polissages arrondis dans le cas du transport par eau. Mais plus que leur origine, c'est la qualité des limons qui intéresse le pédologue, et celle-ci est extrêmement variable. On n'a encore jamais trouvé en Franche-Comté de limons carbonates. Ils sont donc probablement acides à l'origine et plus ou moins appauvris selon la durée du lessivage auquel ils ont été soumis.
o Les épandages de chailles
Les résidus des calcaires à chailles de l'Argovien sont des formations particulièrement mobiles. Ils constituent des boues limono-argileuses et caillouteuses qui se sont beaucoup déplacées au cours du temps. Ramollies par les alternances de gel et de dégel, elles ont glissé, même sur des pentes très faibles. Soumises à l'érosion de leurs particules fines, elles se sont parfois enrichies relativement en chailles. Il en résulte que certains placages sont constitués par une accumulation de ces cailloux siliceux cassés, au cortex patiné par le transport. La pauvreté du substrat est alors aggravée par la compacité de cette formation.
Caractères et distribution des sols de la forêt de Chailluz
La forêt de Chailluz s'étend sur un plateau et les pentes faibles, les actions pédogéniques s'exercent surtout dans les altérites anciennes acides, extrêmement peu influencées par le calcaire, même si ce dernier se trouve proche de la surface ; les sols ont une longue histoire et sont polygéniques.
o Le paradoxe des calcaires durs : des roches-mères acides
Le socle calcaire qui s'étend sur de vastes surfaces porte des sols qui évoluent, de façon totalement indépendante de ce substratum, dans des altérites tertiaires acides.
Dans le Bois des Épesses par exemple (Montfaucon, Doubs), on rencontre des profils de vingt-cinq centimètres formés d'un limon ocre recouvrant un banc massif de calcaire bathonien. On s'attend à un sol neutre ou faiblement acide, bien pourvu en calcium : or, le pH est de 5 et le taux de saturation[4] de 24% !
Ce cas, loin d'être isolé, constitue plutôt la règle. Beaucoup de sols forestiers ont des pH de 4 dans l'humus et sont acides dans la plus grande partie de leur profil (Forêt de Chailluz). On comprend combien la présence du socle calcaire peut être trompeuse. Que ce dernier soit recouvert par des apports allochtones ou par ses propres résidus totalement décarbonatés, il intervient très faiblement dans la pédogenèse. La végétation naturelle en forêt n'apparaît d'ailleurs calcicole que dans des zones très restreintes. Le plus souvent elle traduit les propriétés mésotrophes, voire oligotrophes, des humus et des sols.
o Le caractère polygénique des sols
La longue histoire des pédogenèses successives est inscrite dans les sols. Le meilleur moyen de la reconstituer est d'étudier les microstructures des différents horizons ce qui permet de reconnaître toutes les étapes qui ont façonné le sol actuel. C'est ainsi que l'on retrouve à peu près partout au contact des bancs calcaires, des argiles de décarbonatation. Elles constituent les matériaux dans lesquels les sols ont pris naissance. Les effets des actions périglaciaires s'y manifestent par la présence de nodules arrondis formés par cryoturbation. Mais les argiles de décarbonatation ont le plus souvent été remaniées et érodées. Ce qu'il en reste a été recouvert par plusieurs générations de limons déposés au cours des périodes interglaciaires. La micromorphologie permet là aussi de détecter très nettement ces apports successifs. Enfin, on retrouve la trace de tous les processus d'évolution, en particulier des phases successives de lessivage des argiles, témoins du caractère polygénique des sols.
o L'évolution générale : brunification et lessivage
La plupart des sols de cette région, bien que débarrassés du calcaire, sont tenus à l'abri d'une acidification extrême par la qualité de leurs matériaux minéraux. Il s'agit en effet, à quelques exceptions près que nous signalerons plus loin, de limons toujours en mélange avec des argiles. Or ces dernières par leurs propriétés de colloïdes jouent un rôle fondamental dans la fixation des ions minéraux et la structuration. Elles sont par là un facteur important d'équilibre et de fertilité.
Sur ces roches-mères effectives des sols, l'action du climat aboutit à la "brunification", processus pédogénique dans lequel l'altération des minéraux demeure modérée et le fer prend un état cristallisé caractéristique (goethite), responsable de la couleur brune.
À pH modérément acide, les argiles ont tendance à se mettre en suspension dans l'eau de pluie qui traverse le profil. Elles migrent et se déposent généralement un peu plus bas. Ce transport s'appelle "le lessivage" et conduit à différencier le sol en trois horizons typiques :
A1, organo-minéral grumeleux et teinté par les composés humiques ;
A2, plus minéral et éclairci par perte d'argiles, de fer et d'éléments nutritifs ;
Bt , coloré en brun rougeâtre par le fer associé aux argiles qui revêtent les unités structurales polyédriques (argilanes).
o Les excès de l'évolution : l'hydromorphie et la podzolisation secondaires
La brunification et le lessivage mènent aux profils peu différenciés et peu humifères des sols bruns et bruns lessivés caractéristiques de notre climat atlantique tempéré. Mais avec l'âge, le lessivage s'accentue et conduit aux sols lessivés. Le processus poussé à l'extrême tend à acidifier fortement la partie supérieure des profils (pH de 4 en A1 et A2) et à colmater les pores de la partie inférieure (les argiles entraînées s'accumulent en Bt). Selon les cas, les sols lessivés se dégradent en sols à pseudogley ou bien en sols podzoliques. La première voie s'observe dans les matériaux les plus sensibles à la battance et à la dispersion rapide des argiles, la seconde dans les matériaux capables par contre de conserver une grande perméabilité.
1. Les sols lessivés à pseudogley
Dans les limons très battants, les argiles dispersées dans les horizons supérieurs se déposent et ferment très vite les vides des horizons inférieurs. L'eau ralentit son infiltration puis marque un temps d'arrêt avant d'être évacuée à la fois par le fond, par écoulement latéral et par pompage dû à la végétation. Mais ces mécanismes ne sont pas synchronisés : quand il pleut beaucoup l'hiver, la végétation n'évapore pas et l'eau s'installe pour un certain temps sur le plancher imperméable, constituant ainsi une nappe temporaire.
Au niveau du sol, cela se marque par des transformations de l'état du fer. Dans l'eau stagnante de la nappe, il prend une teinte gris-bleu (état réduit ou ferreux). Quand l'eau se retire, les parties les plus oxydées (trajets des racines, fentes) prennent une teinte rouille (état oxydé ou ferrique). La partie du sol soumise à la présence de la nappe se caractérise par un bariolage de zones grises et rouilles, qui persistent même en période sèche et permettent de diagnostiquer à coup sûr l'hydromophie. Survenant à la suite du lessivage, cette hydromorphie est dite secondaire. Le sol temporairement engorgé est appelé lessivé à pseudogley.
Pour la végétation, cette dégradation est évidemment peu favorable. La zone colmatée fait obstacle à la pénétration des racines. Ces dernières ne prospectent plus qu'un volume réduit de terre. En été, la réserve hydrique s'épuise très vite. En forêt, la végétation naturelle évolue vers des groupements d'espèces capables de supporter alternativement l'hydromorphie asphyxiante et la sécheresse, comme certaines laîches et la molinie.
2. Les sols lessivés podzoliques
Dans les limons grossiers filtrants, l'acidification s'intensifie rapidement dans les horizons supérieurs lessivés. L'activité des microorganismes diminue par manque d'éléments nutritifs. Les débris végétaux se décomposent incomplètement et produisent des acides organiques capables d'agresser et de détruire les minéraux à l'exception du quartz. Ce dernier donne un résidu blanchi, totalement inerte et infertile. Parmi les éléments lessivés, le fer migre avec des matières humiques à l'état de complexes et va précipiter plus bas. Ces mécanismes constituent la dégradation podzolique.
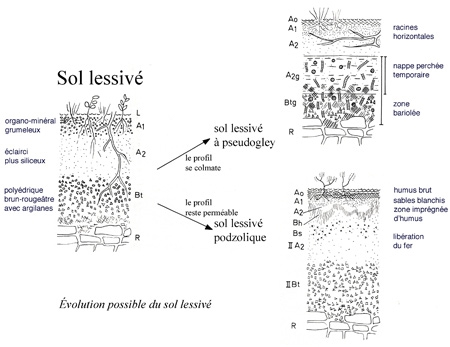
Dans le profil, cela se traduit par plusieurs caractères : la litière s'accumule en couche épaisse et fibreuse (humus brut) ; les grains de silice, dans l'humus et au-dessous sont décapés et apparaissent en blanc sur fond noir (horizons A1 et A2) ; sous une bande un peu éclaircie on voit une couche irrégulière correspondant à l'accumulation de produits organiques brun-rougeâtre qui enrobent et colorent les grains de sable (horizon Bh) ; enfin on découvre un horizon ocre vif ((Bfe), témoin de la forte altération des minéraux et de la libération intense du fer.

De remarquables turricules tricolores qui extériorisent les horizons brun foncé, beige clair et ocre rosé du sol lessivé podzolique sous-jacent. Fontaine Sainte-Agathe, Bois de Chailluz, Doubs (cliché S. Bruckert).
Pour la végétation, la dégradation podzolique est extrêmement préjudiciable. L'humus brut freine en effet le recyclage des constituants absorbés par les plantes. Il bloque les éléments nutritifs sous une forme inassimilable. De tels milieux ne peuvent être colonisés que par des espèces végétales peu exigeantes : la callune, le polytric...
Épaisseur des sols de la forêt de Chailluz
Au cours de sa thèse consacrée à la Forêt de Chailluz, Catherine Fruchart (2014) a pu estimer l'épaisseur des sols forestiers grâce aux techniques LiDAR.
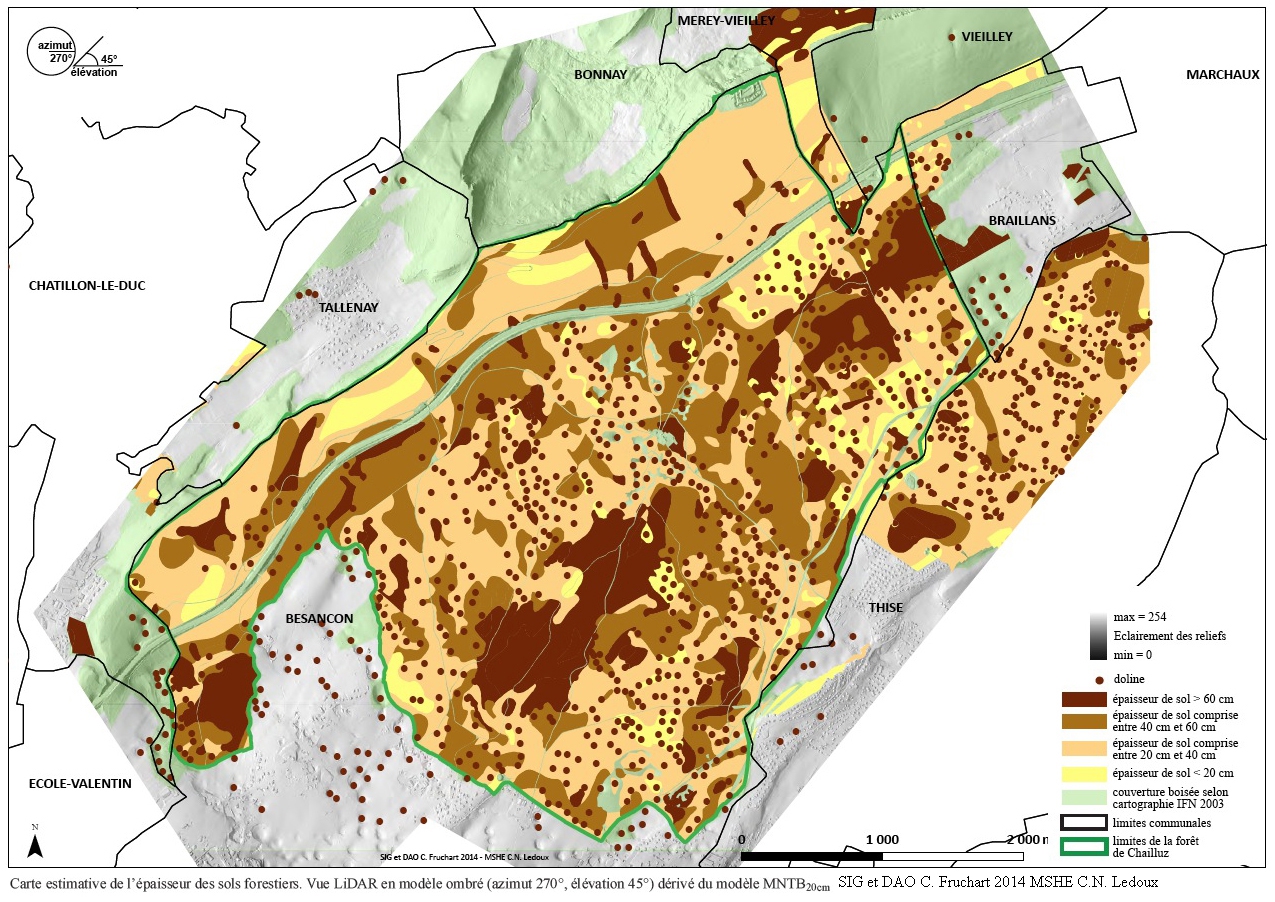
Carte estimative de l'épaisseur des sols forestiers.
Vue LiDAR en modèle ombré
(SIG et DAO C. Fruchart 2014 MSHE C. N. Ledoux)
Valeur pédologique et stationnelle des sols forestiers
Comme pour les terres cultivées, le classement des sols forestiers tient compte de l'estimation analytique du profil pédologique, des conditions stationnelles et du comportement des essences forestières en place. Les propriétés du matériau et la profondeur du sol constituent dans ce cas aussi des critères de qualité. En revanche, les cailloux et les pentes qui entravent les travaux agricoles, les blocs rocheux qui rendent impossible la mise en culture, ne sont plus ici des contraintes ; ce qui compte au plus haut point, c'est le volume de terre meuble prospecté par les racines : les éboulis et colluviums des pentes donnent naissance la plupart du temps à d'excellents sols forestiers, alors que les bancs rocheux compacts n'offrant qu'une mince couche de terre, ont des potentialités forestières médiocres.
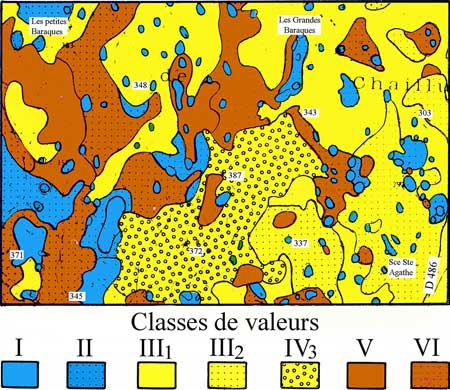
Classes de valeurs des sols de la forêt de Chailluz (d'après S. Bruckert et M. Gaiffe 1985)
Comme dans l'estimation de la valeur des terres agricoles, la démarche méthodologique employée pour réaliser le document présenté vise à classer les sols comme des objets définis par des propriétés précises. L'échelle des valeurs retenue reste totalement indépendante de l'aptitude des sols à cultiver une essence donnée. C'est au forestier que revient ce choix, compte tenu des exigences édaphiques et climatiques de l'espèce, des conditions de sylviculture et d'exploitation, du contexte économique.
L'ENRÉSINEMENT EST-IL UN FACTEUR DE DÉGRADATION DES SOLS FRANCS-COMTOIS ?
On admet généralement que les aiguilles de résineux se décomposent plus difficilement que les litières de feuillus. C'est une des raisons qui font mettre au pilori les résineux, souvent accusés de dégrader les sols, c'est-à-dire d'acidifier l'humus et de provoquer un appauvrissement en éléments nutritifs par lessivage. Dans les cas extrêmes, on assimile cette dégradation à de la podzolisation. Qu'en est-il exactement ? Les résineux et les feuillus ont-ils des actions fondamentalement différentes ? Existe-t-il des sols plus sensibles que d'autres à la dégradation ? Les résineux dégradent-ils les sols ?
De nombreuses observations faites depuis une dizaine d'années démontrent que les résineux sont loin d'avoir tous la même influence nocive : si l'épicéa est en effet très acidifiant, le sapin et surtout le Douglas, le sont beaucoup moins. Quant aux feuillus, leur action peut être tout aussi dégradante ; le hêtre, le chêne ou le châtaignier fournissent des litières qui résistent longuement aux attaques des microorganismes.
On estime maintenant qu'une des premières causes de dégradation est avant tout liée à la monoculture d'une essence forestière, pratique qui se solde inévitablement par une inappétence de la microflore. C'est pour cette raison que les forestiers conservent de plus en plus dans les peuplements un pourcentage de 5 à 15% d'essences secondaires, dans le seul but de maintenir un bon niveau d'activité microbiologique dans l'humus.
Une deuxième cause de dégradation est à rechercher dans les sols. Les matériaux siliceux limoneux ou limono-sableux y sont particulièrement sensibles parce qu'ils ont une faible capacité d'échange cationique et sont inaptes à retenir les cations. Ils sont donc fortement prédisposés à l'acidification.
Pour illustrer ce propos, voici deux exemples choisis au Bois de Chailluz :
|
|
Sols à haut risques (Combottes) |
Sols sans risques (Montarmots) |
|||||||
|
Horizon |
Profondeur en cm |
Argile* |
CEC** |
Taux de saturation.* |
pH |
Argile |
CEC |
Taux de saturation.* |
pH |
|
A1 |
10 |
15 |
10 |
4.4 |
4.1 |
29 |
25 |
63 |
4.9 |
|
A2 |
40 |
16 |
7 |
1.2 |
4.3 |
31 |
25 |
19 |
3.9 |
|
Bt |
60 |
38 |
14 |
2.1 |
4.4 |
47 |
31 |
60 |
5.1 |
|
IIBt |
90 |
26 |
8 |
2.1 |
4.4 |
63 |
33 |
85 |
6.0 |
* en % ** en milliéquivalents pour 100 grammes
Le sol des Combottes qui s'est forme aux dépens d'un faciès gréseux de l'Argovien se distingue par une capacité d'échange et un taux de saturation en cations extrêmement bas : ses caractéristiques analytiques le font ranger dans les sols à hauts risques. En revanche, celui des Montarmots développé dans un limon de meilleure qualité possède une capacité d'échange et un taux de saturation beaucoup plus élevés qui les rangent dans les sols sans risques.
Ces observations répétées sur un grand nombre d'échantillons, démontrent que ce sont effectivement ces deux données analytiques qui permettent de diagnostiquer d'une façon simple et sûre la sensibilité à la dégradation. On soulignera au passage que le pH des 40 premiers centimètres ne distingue absolument pas les deux sols.
[1] Les pédologues dénomment "horizons" les strates successives de couleur, texture et structure différentes, dont l'ensemble constitue le profil d'un sol, mis à jour par une coupe verticale.
[2] Compte tenu de la toxicité de la plupart des pesticides pour les lombrics, on est en droit de s'interroger sur les conséquences de leur usage, à long terme, sur la fertilité des terres cultivées.
[3] Rappelons que l'on appelle limons, les particules dont la taille est comprise entre 2 et 50 millièmes de millimètre.
[4] Taux de saturation : proportion des sites effectivement occupés par les cations Ça2+ , Mg2+, K+, par rapport aux sites électro-négatifs potentiels : les sites non saturés sont comblés par des ions H+.
Sources :
22:12 Publié dans Botanique, Environnement-Écologie, Géologie-hydrogéologie-Climatologie | Tags : pédologie, franche-comté, forêt de chailluz | Lien permanent | Commentaires (0) | Facebook | | |
08/03/2014
Usage massif des OGM aux États-Unis : pas de réduction des herbicides
Usage massif des OGM aux États-Unis : pas de réduction des herbicides
(Sciences et Avenir n° 806 Avril 2014, p.18)
Le Département américain de l'agriculture vient de publier le bilan de plus de quinze ans de culture d'organismes génétiquement modifiés, les premières semences ayant été commercialisées en 1996. Ce document montre que, si au cours de ces quinze ans, les cultures OGM ont progressé aux États-Unis, seule l'utilisation des insecticides a baissé. En revanche l'usage massif des OGM n'a pas limité les herbicides. En somme, les agriculteurs américains ont adopté ces cultures pour augmenter la production et non pas pour réduire les pesticides.
Certes, le succès des plants génétiquement modifiés auprès des agriculteurs américains est incontestable, mais le bilan est mitigé. Si le coton, le soja et le maïs OGM sont utilisés massivement, c'est en raison de leur rendement supérieur et non pour diminuer le recours aux pesticides.
Si, par ailleurs, la réduction des insecticides est réelle, elle est due principalement à une baisse très importante des populations de pyrales, des papillons ravageurs du maïs et d'autres plantes cultivées, laquelle profite aussi aux cultures non OGM. De plus, la création de zones non OGM a permis de limiter l'émergence de résistances aux pesticides chez les insectes.
En revanche, des résistances aux herbicides sont apparues pour 14 "mauvaises herbes", ce qui a contraint les agriculteurs à maintenir les épandages chimiques.
15:55 Publié dans Actualité des Sciences, Botanique, Environnement-Écologie, Pollution | Tags : ogm, pesticides, insecticides, herbicides | Lien permanent | Commentaires (0) | Facebook | | |
10/02/2014
Le Printemps des Campenottes
Le Printemps des Campenottes
par Dominique Delfino
Photographe naturaliste et animalier
Oui elle est là bien là, illuminant les sous-bois de sa couleur jaune vif pour nous rappeler qu'elle est avant tout la fleur du printemps.
La Campenotte, comme on la surnomme dans le Pays de Montbéliard est chère à ses habitants. Une histoire ou légende de vieux Druide et sa fille vivant dans le Mont-Bart lui accorderait l'origine de sa naissance à cet endroit magique.
Alors quelle qu’en soit la croyance, les centaines de jonquilles qui tapissent le sol sont toutes autant de raisons de poser le regard sur chacune d’entre elles, la lumière qui les traverse, transformant chaque corolle en une petite tache couleur d'or.

Cliché © Dominique Delfino
La Jonquille (Narcissus pseudonarcissus, famille des Amaryllidaceae), appelée Campenotte en Franche-Comté a fait l'objet d'une chanson que publie le site "La Cancoillotte".
Sur l’air de "Ma Normandie"
Premier couplet
Dans Paris, fleur abandonnée,
De mon pays doux souvenir.
Ô ! Campenotte infortunée
Toi qui viens jusqu’ici mourir,
N’as-tu pas vu dans ton voyage
Ma jolie diaichotte aux yeux doux ?
Ô ! parle moi de mon village,
De mon pays et des rives du Doubs.
Deuxième couplet
Si tu viens de cette montagne
Qui de l’Allan voit les roseaux,
Si tu as vu notre campagne
Du haut de la roche aux corbeaux,
Ah ! tu dois connaître je gage
Ma jolie diaichotte aux yeux doux,
C’est la plus belle du village
De mon pays et des rives du Doubs.
Troisième couplet
As-tu vu passer le dimanche
En corset noir baissant les yeux,
Les bras nus sous la guimpe blanche
Le diairi sur ses longs cheveux
Rubans flottants sur son corsage,
Ma jolie diaichotte aux yeux doux ?
C’est la plus belle du village,
De mon pays et des rives du Doubs.
Quatrième couplet
Mais si ma mie est la plus belle
Toi, notre emblème de gaieté,
Vous deux à la saison nouvelle
Serez toujours de la Comté
Toi, reine de nos fleurs sauvage
Et la diaichotte aux yeux si doux,
Reine des fleurs et des villages
De mon pays et des rives du Doubs.
Paroles de P. de Resener, dédié au poète Frédéric Bataille et chantée pour la première fois le 11 juillet 1891 à Montbéliard par M. Jordy.
14:58 Publié dans Art et Nature, Botanique, Nature & philosophie, Photographes naturalistes et scientifiques | Lien permanent | Commentaires (0) | Facebook | | |
08/02/2014
Massacre de haies dans le pays de Montbéliard
Pas de sentiments pour les haies
par Dominique Delfino
Photographe naturaliste et animalier
C'est un véritable massacre végétal que l'on peut constater dans la pâture qui borde l'A36 avant la Savoureuse sur la commune de Vieux-Charmont.

À mon retour de voyage, je constate que l'ensemble des haies et massifs buissonnants qui caractérisaient le biotope de cette pâture, avaient été coupés. En me rendant sur place, le spectacle de désolation est à son comble. Ce sont près d'une centaine de massifs qui ont été arrachés, déracinés, broyés, laissant place nette, seuls les pylônes électriques sont encore debout, mais il s'en fallait de peu !
Le service environnement de PMA me confirme qu'il avait également constaté cet état de faits mais que les terrains sont totalement privés sans contrôle de leur services.

Alors qu'aujourd'hui, l'intérêt majeur que représente les haies n'est plus à démontrer, ce type de gestion radicale d'un milieu naturel est grave de conséquences pour la faune locale surtout dans un secteur au contact de l'agglomération montbéliardaise.
La Pie Grièche-écorcheur, espèce protégée dont les effectifs se font de plus en plus rares, trouvait là le biotope idéal. Il est dorénavant sûr de ne plus la revoir comme d'ailleurs d'autres espèces dépendantes de ces milieux ouverts clairsemés d'espaces buissonnants devenus rares sur le Pays de Montbéliard.

Un très bon ami naturaliste me confiait, il y a quelques années que les hivers doux étaient souvent fatals pour la nature, car certains agriculteurs passaient leur temps à couper les vieux arbres et les haies à tort et à travers... La preuve en est.
07:37 Publié dans Botanique, Environnement-Écologie, Nature et santé, Photographes naturalistes et scientifiques | Tags : dominique delfino, destruction de haies, pays de montbéliard, vieux-charmonthaies | Lien permanent | Commentaires (1) | Facebook | | |
04/11/2013
Stage de permaculture à Froidevaux
Stage de permaculture à Froidevaux
Ce stage de permaculture est co-organisé par une nouvelle association créée pour l'occasion (en fait le potager d'une curieuse avec quelques amis) et Bernard Alonso, permaculteur canadien qui est déjà venu à Besançon en 2012.
La permaculture est un ensemble de pratiques et de modes de pensée visant à créer une production agricole soutenable, très économe en énergie (travail manuel et mécanique, carburant...) et respectueuse des êtres vivants et de leurs relations réciproques. Elle vise à créer un écosystème productif en nourriture ainsi qu'en d'autres ressources utiles, tout en laissant à la nature "sauvage" le plus de place possible.
Elle utilise des notions d'écologie, de paysagisme, d'agriculture biologique, de biomimétisme, d'éthique, de philosophie et de pédologie. La permaculture invite à mettre ces aspects théoriques en relation avec les observations réalisées sur le terrain.
La base de la permaculture n'est pas uniquement d'analyser les éléments constitutifs d'un système individuellement, mais aussi de prendre en compte leurs interactions, dans le but de produire une compréhension de l'écosystème dans l'optique d'une utilisation par l'Homme.



08:13 Publié dans Biologie, Botanique, Environnement-Écologie, Nature & philosophie, Nature et santé | Lien permanent | Commentaires (0) | Facebook | | |
11/04/2013
Le temps des bourgeons
Le temps des bourgeons
par Dominique Delfino
photographe animalier
Le printemps est bien là, même si les températures de ces derniers jours nous font encore douter. La nature a néanmoins profité des journées très ensoleillées de la semaine dernière favorisant l'éclosion spontanée des bourgeons et des feuilles en forêt.
Très friands de cette végétation qui s'offre à eux après l'hiver, les chevreuils trouveront là une réserve de nourriture presque inépuisable.
Chaque année, au printemps, des chevreuils se retrouvent en ville après avoir consommé des bourgeons gorgés de sève qui les enivrent en fermentant dans leur estomac. Désorientés, titubants, ils se trompent alors de chemin…
Le cliché que j'ai pu réaliser lors de cette rencontre avec ce brocard en forêt témoigne de l'intérêt que celui-ci manifeste pour ces feuilles tendres qu'il consommera jusqu'à les cueillir au plus haut des branches sans trop se préoccuper de ma présence discrète.

Cliché © Dominique Delfino
22:35 Publié dans Botanique, Environnement-Écologie, Mammifères, Photographes naturalistes et scientifiques | Tags : chevreuil, dominique delfino, printemps, bourgeons, ivresse | Lien permanent | Commentaires (0) | Facebook | | |
19/02/2013
Le Callitriche des étangs
Le Callitriche des étangs
Le Callitriche des étangs ou Étoile d'eau [Callitriche stagnalis SCOP] appartient à la famille des Callitrichacées.
La plante se présente comme un semis d'étoiles. Les Callitriches sont des hydrophytes fixés et dont le feuillage présente un aspect artistique.
Le classement systématique des Callitriches et la recherche de liens de parenté avec d'autres plantes sont très difficiles. L'ordre des Callitrichales est formé d'une seule famille les Callitrichacées qui ne comportent que le seul genre Callitriche. Ces plantes, nées sous le signe de l'unicité, ont des fleurs unisexuées et les fleurs mâles sont réduites à une seule étamine.
Ce sont des plantes très variables, petites et fragiles, qui ont des formes aquatiques, immergées ou flottantes, et des formes terrestres, qui vivent, éphémères, sur la boue humide.
Le Callitriche des étangs (C. stagnalis) pousse le plus souvent dans les eaux peu profondes, dormantes ou à faible courant ; plus rarement sur un sol humide. En Europe, on le trouve surtout dans l'ouest et le centre ; C. palustris L. est plus abondant ; il est considéré comme un élément arctico-alpin qui pousse même en Scandinavie et en Islande. En Angleterre, on ne le connaît que dans une seule station. Il supporte bien le froid et verdoie parfois sous la glace.

Les Callitriches sont des plantes vivaces quand elles poussent dans l'eau, annuelles quand elles poussent sur terre, fines, vert vif, à feuilles étroitement linéaires, groupées en étoiles au sommet de la plante, particulièrement remarquables quand on les voit de haut (1).
Les fleurs unisexuées sont très réduites ; les fleurs mâles n'ont qu'une seule étamine, les fleurs femelles un seul pistil à 2 styles et 4 ovules. Les pièces florales se réduisent à 2 écailles. Les fruits de C. stagnalis (2) sont entourés par des ailes et les styles, s'ils se maintiennent, sont retombants. Les fruits de C. palustris (3) ont des ailes peu développées à leur partie supérieure et les styles disparaissent rapidement.
Les fleurs s'épanouissent l'une après l'autre du mois d'avril au mois d'octobre.
Source :
V. Vetvicka (1981). - Plantes du bord de l'eau et des prairies. Ed. Gründ.
09:14 Publié dans Botanique, Limnologie-hydrobiologie | Tags : callitriche, étoile d'eau, zones humides, hydrophytes | Lien permanent | Commentaires (0) | Facebook | | |
Les Lentilles d'eau
Les Lentilles d'eau
Les Lentilles d'eau appartiennent à la famille des Lemnacées. Ce sont de petites plantes aquatiques flottant à la surface dont les organes sont très réduits. Leur corps est représenté par un organe foliaire considéré soit comme une seule vraie feuille plus ou moins transformée, soit comme une tige plate en forme de feuille. Dans le genre Spirodela, on peut dire que la grande feuille plate est une tige car elle possède en dessous deux vraies feuilles en écailles. Les Lentilles d'eau fleurissent très rarement, mais se multiplient végétativement de façon très intense. Dans la "feuille" maternelle se trouve un creux qui contient la gemmule : elle donne une nouvelle "feuille" qui s'émancipe chez la plupart des Lentilles d'eau mais reste attachée chez certaines autres (Lemna trisuica).
Les Lentilles d'eau sont des plantes cosmopolites transportées facilement et à de grandes distances, par exemple sur les pattes et dans les plumes des oiseaux aquatiques. Elles poussent dans toutes les eaux calmes, souvent en très grand nombre. La représentation des différentes espèces dans leurs associations peut varier considérablement au cours d'une même période de végétation : c'est ainsi que dans un étang, entre le mois de mai et le mois de juillet, on a constaté une extension continue de Spirodela polyrhiza (L) SCHL. suivie de son déclin. Lemna trisulca évolua de manière inverse, tandis que Lemna minor déclina continûment depuis son maximum printanier.

La Petite Lentille d'eau (Lemna minor L.) a des feuilles (articles) plates aux deux faces vertes, flottant à la surface, et une seule racine (1).
La Lenticule bossue (Lemna gibba L) a des feuilles gonflées (2).
La Lenticule à trois lobes (Lemna trisuica L ) a des feuilles ovoïdes, lancéolées, comme enfoncées dans l'eau, à longs pétioles et attachées ensemble (3) pendant plusieurs "générations".
La Spirodèle à racines nombreuses [Spirodela polyrhiza (L) SCHL] a des feuilles ovoïdes arrondies à l'envers rougeâtre et portant chacune un petit faisceau de racines filamenteuses (4).
Les fleurs des Lentilles d'eau, quand elles fleurissent, sont réduites à une seule étamine ou un seul pistil.
Source :
V. Vetvicka (1981). - Plantes du bord de l'eau et des prairies. Ed. Gründ.
09:07 Publié dans Botanique, Limnologie-hydrobiologie | Tags : lentilles d'eau, lemnacées, zones humides, hydrophytes | Lien permanent | Commentaires (0) | Facebook | | |
Les Renoncules d'eau
La Renoncule flottante
La Renoncule flottante [Ranunculus fluitans Lam., 1779] appartient à la famille des Renonculacées.
Les surfaces aquatiques où fleurissent des Renoncules d'eau ont l'air d'être couvertes d'une sorte d'écume blanchâtre. Et l'impression produite par la masse fleurie des Renoncules est renforcée du fait qu'elles ne sont pas seules dans les eaux calmes.
La Renoncule flottante (Ranunculus fluitans) habite les cours d'eau au plus fort du courant Dans les zones calmes où elles sont enracinées, elles forment des amas épais entraînés par les eaux où s'abritent souvent les brochets et les anguilles.
La Renoncule aquatique [Ranunculus aquatilis (L.) DUM.] plante importante, pratiquement cosmopolite, préfère les eaux dormantes, les mares et les bras morts des cours d'eau, à 50 cm de profondeur environ. Sur les fonds asséchés, et sur les rives, elle produit une forme terrestre aux feuilles entières.
Les Renoncules d'eau sont souvent classées dans le genre Ranunculus. De récentes études sur l'anatomie, la morphologie et la biologie florales de ces plantes montrent qu'elles forment un groupe séparé, indépendant. Les Renoncules d'eau sont des plantes vivaces dépendant presque entièrement du milieu aquatique. Les feuilles submergées des plantes flottantes sont souvent divisées en filaments.

Les plantes et les filaments foliaires de Ranunculus fluitans (1) flottent avec le courant ; les feuilles de Ranunculus aquatilis sont doubles : feuilles immergées en filaments et feuilles flottantes ou émergées dont le limbe est plat (2).
Les fleurs sont à 5 divisions (3) ; mais il y a parfois jusqu'à 7 pétales. Les Renoncules qui fleurissent en juin ont des fleurs beaucoup plus grandes que celles qui refleurissent en août et septembre.
Les fleurs fécondées donnent une tête (4) composée d'akènes bien séparés (5) qui flottent à la surface.
Source :
V. Vetvicka (1981). - Plantes du bord de l'eau et des prairies. Ed. Gründ.
09:02 Publié dans Botanique, Limnologie-hydrobiologie | Tags : renoncule aquatique, renoncule flottante, zones humides, hydrophytes | Lien permanent | Commentaires (0) | Facebook | | |