La différenciation des gamètes dans le sens mâle ou dans le sens femelle est sans doute due à des interactions hormonales. Il faut donc rechercher des organes endocrines susceptibles d'agir sur la gamétogenèse.
 L'Escargot (11) : Organes présumés endocrines
L'Escargot (11) : Organes présumés endocrines
par André Guyard
(suite de l'Escargot 10)
Chez les Mollusques, on a décrit des organes à fonction endocrine "possible" et l'existence de phénomènes neurosécrétoires peut être considérée comme "probable.
Parmi les organes susceptibles d'avoir une action sur la différenciation des gamètes qu'on a testés, on n'envisagera ici que les organes suivants :
- les cellules neurosécrétrices du cerveau, c'est-à-dire des ganglions cérébroïdes ;
- les cellules neurosécrétrices situées dans le ganglion piriforme des tentacules oculaires.
En ce qui concerne la gonade elle-même, la fonction sécrétoire éventuelle peut se localiser à deux niveaux :
- au niveau des cellules nourricières dans lesquelles l'étude ultrastructurale révèle des inclusions ;
- au niveau du tissu conjonctif intertubulaire.
Chez l'Escargot, le développement de ce tissu périgonadique précède la mise en place de l'ovogenèse. Dernier intermédiaire entre l'organisme et l'épithélium germinatif, il participe obligatoirement à tous les échanges humoraux pouvant intéresser la gonade. Il peut être considéré comme le réservoir neurohémal ultime, dernier relais qui collationne les messages humoraux provenant du milieu intérieur et du tissu nerveux à destination de la gonade. Lors de la culture organotypique, il ne peut être dissocié de la gonade.
D'autre part, il faut montrer qu'il y a synchronisme entre cycle sécrétoire des organes endocrines et cycle reproducteur.
Les ganglions cérébroïdes
Les ganglions cérébroïdes constituent la partie dorsale du collier nerveux périœsophagien. On peut les considérer comme le "cerveau" de l'Escargot.
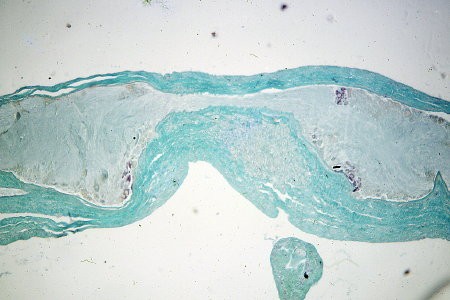
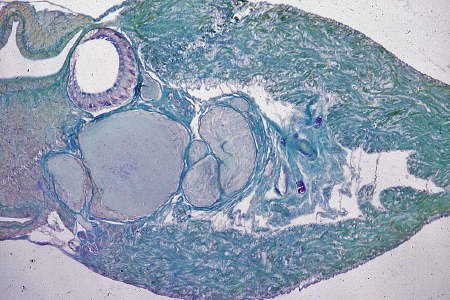
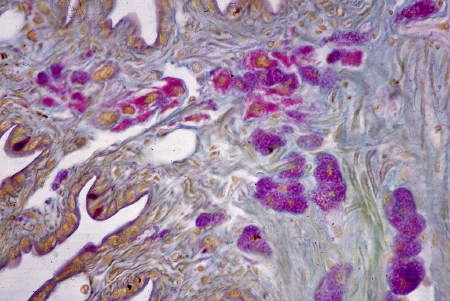
Coupe histologique des ganglions cérébroïdes
Quatre groupes de cellules neurosécrétrices
de type B apparaissent en rose
Chez l'Escargot, la gaine conjonctive (ici colorée en vert foncé) entourant le cerveau peut être considéré comme un organe neurohémal à la façon des corps dorsaux des Pulmonés Basommatophores. Comme elle n'est pas dissociée des ganglions lors de la mise en culture, c'est l'ensemble de ces structures histologiques que nous nommerons "cerveau" dans les associations d'organes.
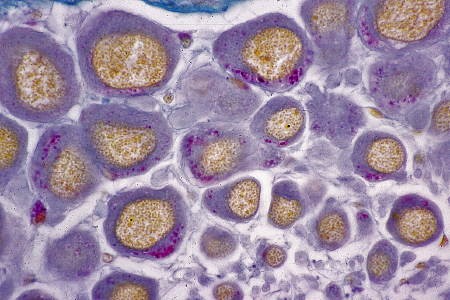
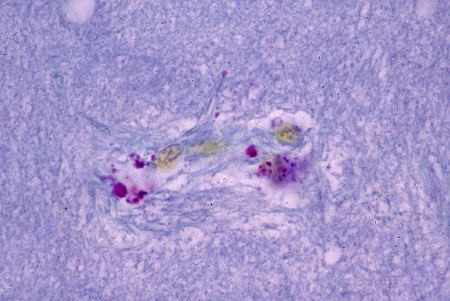
Par les méthodes de la fuchsine-paraldéhyde et de l'hématoxyline chromique-phloxine, on reconnaît deux types cellulaires à fonction neurosécrétoire présumée. Le type A comporte des granules de forte taille, bien visible au microscope photonique (voir la photographie ci-dessous), répandus dans la masse cérébroïde ainsi que dans la masse sous-œsophagienne. Ces lipofuscines seraient étrangères au phénomène neurosécrétoire.
Cellules neurosécrétrice de type A
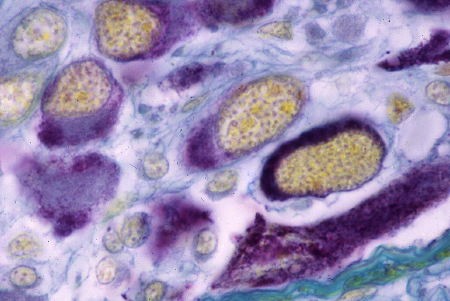
En revanche, la sécrétion des quatre groupes cellulaires localisés dans le cerveau moyen (mésocérébron) se caractérise par des granules ténus inframicroscopiques dont l'abondance forme une masse diffuse qui envahit le péricaryon et s'écoule par l'axone (voir ci-dessous). Les axones des quatre groupes cellulaires se regroupent pour former le nerd de l'artère cérébrale. L'organe neurohémal (réservoir hormonal) est constitué par le tissu conjonctif autour de l'artère cérébrale. La photographie ci-dessous montre les cellules neurosécrétrices de type B qui contiennent des granules petits et abondants formant un voile continu dans le péricaryon et l'axone .
Cellules de type B localisées dans le mésocérébron
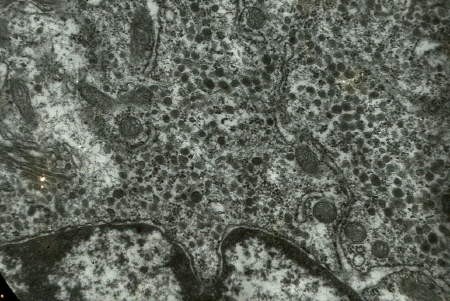
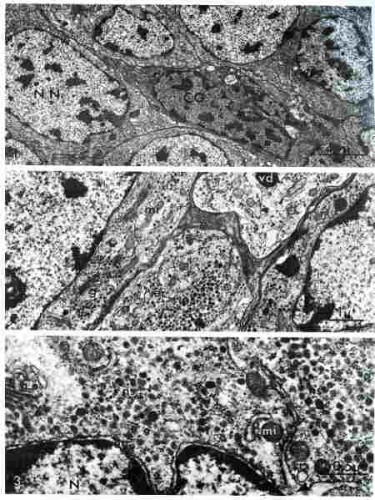
À l'échelon inframicroscopique, presque toutes les cellules du mésocérébron se révèlent riches en grains de neurosécrétion de type B. (voir ci-dessous).Ces grains de neurosécrétion proviennent de la fusion de vésicules golgiennes. En bas, le noyau.
Cellule du mésocérébron
(électronographie)
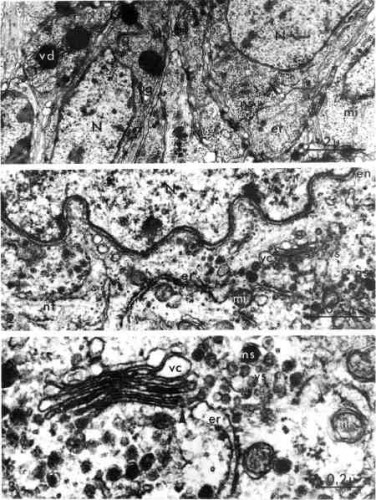
La vue suivante montre une coupe du cerveau au niveau du mésocérébron. ns : granules de neurosécrétion ; N : noyau ; g : dictyosomes ;mi : mitochondries ; en : enveloppe nucléaire ; nf : neurofibrilles ; er : ergastoplasme ; vc : vésicules golgiennes claires ; vs : vésicules golgiennes opaques ;
vd : granules de neurosécrétion.
Coupe au niveau du mésocérébron
(électronographies)
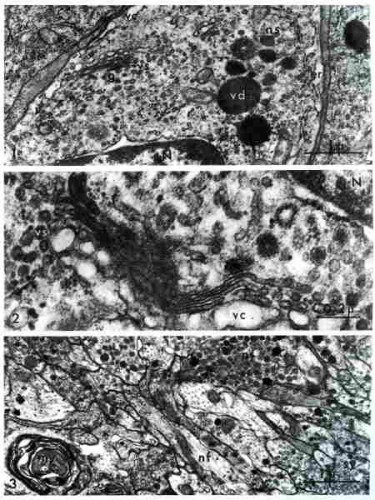
Les trois clichés ci-dessous montrent le cheminement de la neurosécrétion depuis le corps neuronal jusque dans les axones formant les nerfs cérébraux. Le réticulum endoplasmique (er) est constitué de citernes. mi : mitochondries ; nf : neurofibrilles ; ns : vésicules de neurosécrétion (fig. 1). Les grains de neurosécrétion fusionnent (fig. 2). La figure 3 est une coupe longitudinale du nerf cérébral. Le nerf ext constitué d'axones banaux avec neurofibrilles uniquement (nf) et d'axones neurosécréteurs (ns).
Fig. 1 : Cytoplasme d'un neurone
Fig 2 : Coalescence des granules de neurosécrétion
Fig. 3 : Coupe longitudinale du nerf cérébral
(électronographies)
Le ganglion tentaculaire
Dans cette coupe longitudinale du tentacule oculaire (cliché ci-dessous), on distingue l'œil et les structures nerveuses sous-jacentes dont le ganglion digité au centre et à gauche le bouton terminal.
Coupe longitudinale du tentacule oculaire
La microscopie photonique ne révèle, dans le ganglion digité situé sous l'œil qu'un groupe de trois à quatre cellules fuchsinophiles incluses dans le nerf oculaire (neuropile).
Groupe de cellules fuchsinophiles incluses
dans le neuropile du ganglion digité
Dans le bouton terminal, deux types cellulaires répondent à la coloration fuchsine-paraldéhyde : de grosses cellules pourpres et des cellules plus petites d'un rouge carmin. En outre, des cellules muqueuses du tégument prennent également la coloration (voir cliché ci-dessous).
Deux types cellulaires neurosécréteurs sont visibles
dans le bouton terminal
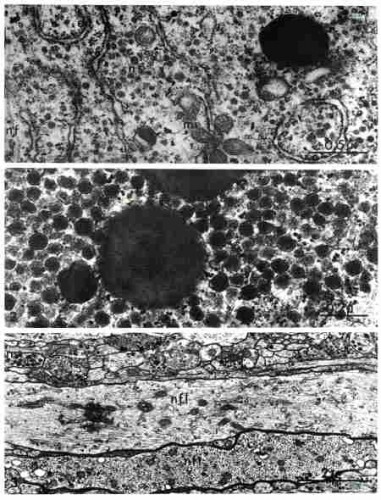
En microscopie électronique, comme dans le cas du ganglion cérébroïde, le péricaryon des neurones du tentacule oculaire se révèle riches en vésicules de neurosécrétion non révélées par la microscopie photonique.
Cellules ganglionnaires dans le tentacule oculaire
NN : noyau des neurones ; CG : cellule gliale
(électronographies)
Les trois électronographies suivantes montrent la neurosécrétion dans le ganglion tentaculaire. vs : vésicules sombres golgiennes ; v : vésicules claires golgiennes ; ns : granules de neurosécrétion ; d : vésicules de grande taille ; N : noyau : face de formation du dictyosome avec vésicules claires (vc) ; face de maturation avec vésicules sombres (vs). nf : neurofibrilles dans axones banaux ; my : fibres myéliniques ; ns : axones neurosécréteurs.
Ganglion tentaculaire. Fig. 1 : Neurosécrétion. Fig. 2 : Dictyosome
Fig. 3 : Transport de la neurosécrétion par voie axonale
(électronographies)
De nombreux chercheurs ont montré un parallélisme entre l'évolution de la neurosécrétion et le cycle sexuel des Mollusques Gastéropodes.
Mais l'existence de ce parallélisme n'autorise pas à conclure à des relations de cause à effet entre cycle neurosécrétoire et cycle sexuel. Si l'on attribue à la neurosécrétion une fonction hormonale, un fort contenu sécrétoire des neurones ne signifie pas nécessairement une importante évacuation de neurosécrétion dans l'hémolymphe. Une telle image peut traduire, au contraire, une forte accumulation avec un vidage réduit. En outre, les coïncidences chronologiques entre cycle neurosécrétoire et cycle sexuel peuvent traduire les réactions simultanées de deux processus physiologiques étranges l'un à l'autre à des stimuli d'origine exogène : variation de la température, de l'hygrométrie, durée ou intensité de l'illumination.
Conclusion :
Le recours à l'expérimentation est nécessaire pour apporter des preuves à l'existence de relations de cause à effet entre l'évolution neurosécrétoire et le cycle sexuel chez les Gastéropodes.
Pour mettre en évidence le rôle présumé d'un organe, l'endocrinologie expérimentale dispose de différentes techniques souvent complémentaires :
- ablation,
- destruction par irradiation,
- implantation,
- injection d'extraits organiques présumés actifs,
- culture organotypique.
C'est cette dernière technique qui permet de mettre en évidence l'action directe d'un organe sur un autre que nous allons éprouver dans les articles suivants qui font partie de cette série.
Sources :
- Guyard A. (1971) - Étude de la différenciation de l'ovotestis et des facteurs contrôlant l'orientation sexuelle des gonocytes de l'Escargot Helix aspersa Müller. Thèse d'État soutenue à la Faculté des Sciences de l'Université de Franche-Comté.
- Documents inédits tirés de la photothèque personnelle de André Guyard.




 Facebook | | |
Facebook | | |
Les commentaires sont fermés.