La genèse de la gonade permet de confirmer que les gamètes des deux sexes proviennent des mêmes cellules souches présentant une morphologie semblable à celle des cellules somatiques.
 L'Escargot (10) : genèse de la gonade
L'Escargot (10) : genèse de la gonade
par André Guyard
(suite de l'Escargot 9 )
Au cours des articles précédents regroupés dans la même série numérotée, nous avons pu supposer que les gonocytes des deux sexes qui apparaissaient dans la glande hermaphrodite provenaient d'une seule et même catégorie de cellules souches.
Afin de vérifier cette hypothèse, une première démarche consistait à observer comment l'ébauche de la gonade apparaissait au cours de l'embryogenèse et la post-embryogenèse de l'Escargot. Nous avons travaillé sur le Petit-Gris, Helix aspersa, dont l'élevage est facile.
Chez les Gastéropodes Pulmonés, l'origine de la gonade est controversée et fait l'objet d'une alternative entre deux théories.
1. La théorie dualiste ou multiciste : deux ébauches au moins collaborent à l'édification de l'appareil génital : l'une ectodermique engendrant le tractus génital et l'autre mésodermique donnant naissance à la gonade et à son canal hermaphrodite.
2. La théorie uniciste : l'ensemble de l'appareil génital, y compris la gonade, provient d'une ébauche unique soit mésodermique, soit ectodermique.
Mes observations personnelles vont dans le sens de cette seconde hypothèse.
L'apparition de la gonade étant contemporaine de l'éclosion chez les Hélicidés, je me suis adressé directement à des animaux postembryonnaires afin de trouver un stade gonocytaire suffisamment précoce pour être sexuellement indifférencié.
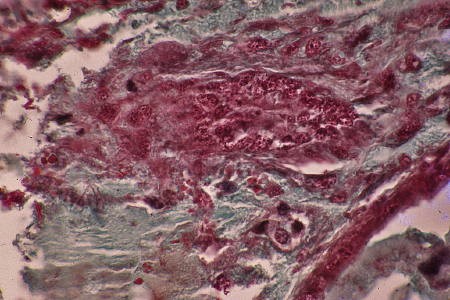
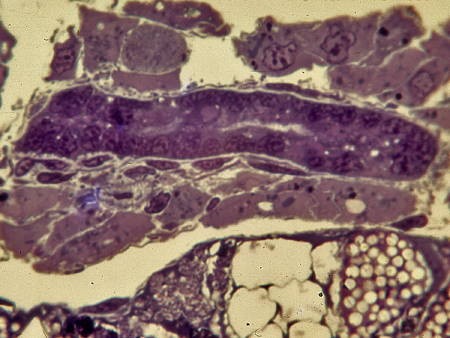
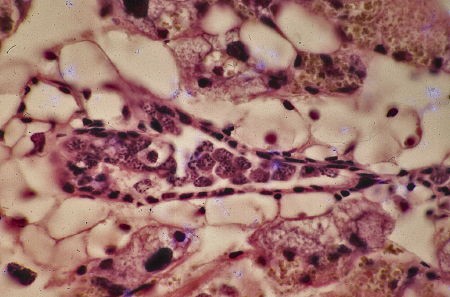
À l'éclosion, l'ébauche gonadique se présente comme un massif cellulaire cylindrique de 25 µm de diamètre sur une centaine de µm de long. J'ai appelé ce stade : stade 1 ou stade du manchon gonadique.
À l'éclosion, l'ébauche gonadique se présente
comme un cylindre massif
La multiplication cellulaire est active
dans l'ébauche gonadique
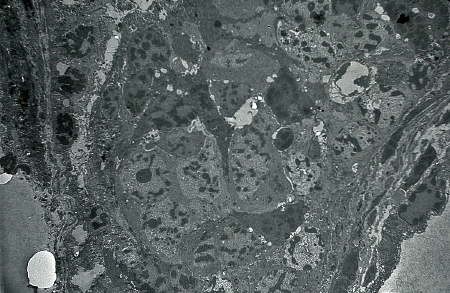
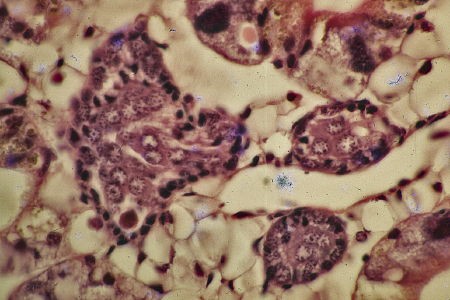
On note une prolifération active des cellules souches distribuées sur la paroi externe du cylindre engendrant des protogonies constituant le massif cellulaire interne. En suivant le devenir de ces protogonies, on s'aperçoit qu'elles auront un destin mâle. C'est le stade 2 ou stade du bourgeon germinal à tendance mâle.
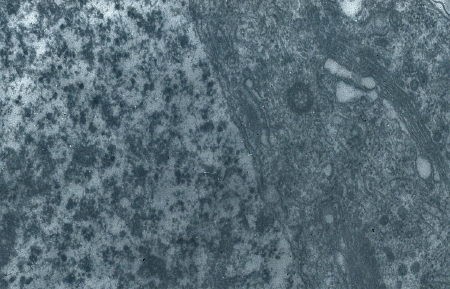
L'ébauche gonadique se présente
comme un cylindre massif
(électronographie)
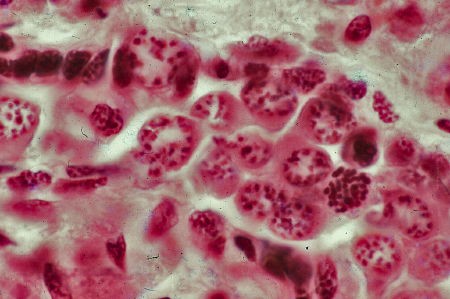
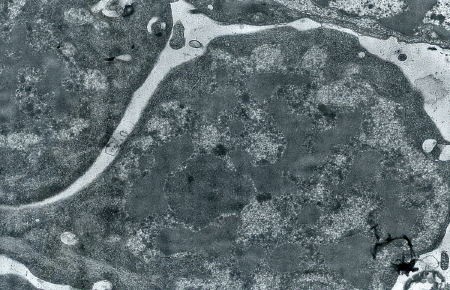
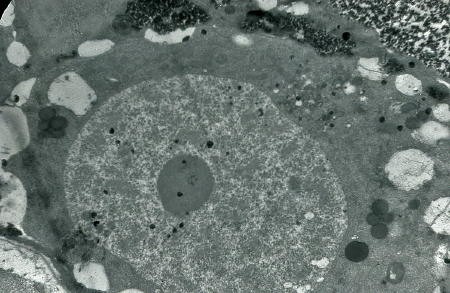
Détail d'une cellule souche à l'éclosion
(électronographie)
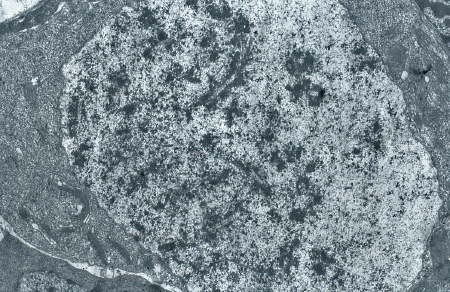
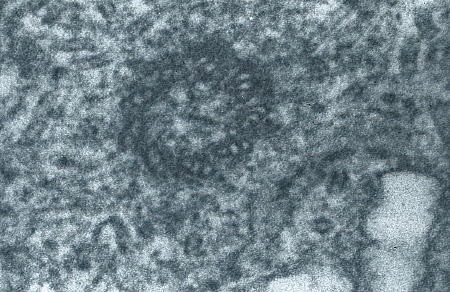
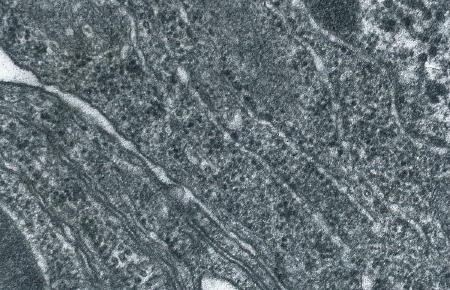
Protogonie en division. Les chromosomes sont visibles
(électronographie)
Détail d'un chromosome dans la protogonie
(électronographie)
Au nord-est du noyau on aperçoit un centriole et
les saccules empilés de l'appareil de Golgi
(électronographie)
Détail du centriole de la protogonie
(électronographie)
Rapidement le cylindre va se creuser en un tubule
(Coupe semi-fine pour la préparation en microscopie électronique)
Dès lors, deux phénomènes vont se combiner : une prolifération des protogonies conjuguée à une dégénérescence de certaines cellules qui vont naturellement se nécroser. Cette mort cellulaire programmée ou apoptose est un phénomène bien connu en embryologie. C'est elle qui préside en particulier à la disparition des palmes existant entre les doigts des fœtus humains. Eh bien, cette apoptose aboutit à l'apparition d'une lumière dans le bourgeon gonadique aboutissant à la formation d'un tubule. Nous atteignons alors le stade 3 ou stade de la gonade cavitaire avec mise en place de l'épithélium germinatif définitif.
Multiplication cellulaire et apoptose cohabitent dans l'ébauche gonadique
(Coupe semi-fine)
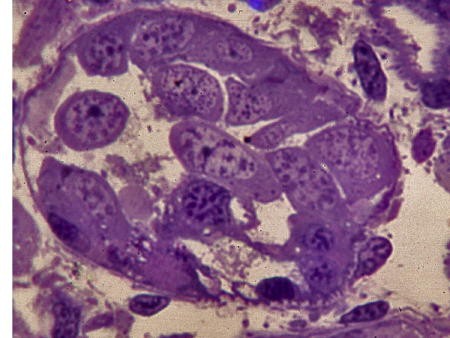
Une dizaine de jours après l'éclosion, le tubule primitif va se ramifier pour engendrer une glande multitubulaire.
Digitation progressive dans la gonade postembryonnaire
L'épithélium germinatif est désormais bien constitué de cellules-souches pariétales bien distinctes des protogonies en situation médullaire vont évoluer en spermatogonies. Comme nous l'avons observé dans le cas de la gonade adulte, la cellule pariétale issue de la division va servir de cellule nourricière au bouquet de spermatogonies médullaires.
Les protogonies évoluent dans le sens mâle et forment un bouquet de spermatogonies
(électronographie)
Détail de deux pédoncules spermatogoniaux montrant
un réticulum endoplasmique riche en ribosomes
(électronographie)
On peut résumer l'évolution postembryonnaire de la gonade par le schéma ci-dessous :
Puis apparaîtront en position pariétale des ovocytes issus de cellules souches. Il est difficile de parler de protogonie femelle puisqu'une cellule femelle ne peut être identifiée que lorsqu'elle est en accroissement donc au stade ovocytaire. L'ovocyte va s'entourer de cellules folliculeuses issues des cellules souches voisines.
On peut donc résumer l'évolution de l'épithélium germinatif dans la gonade postembryonnaire par le schéma suivant :
Ovocyte apparu sur la paroi d'un tubule gonadique
(électronographie)
En conclusion, à partir de cellules souches morphologiquement identiques, l'épithélium germinatif engendre quatre catégories de cellules qui se différencient en deux étapes :
1. Tout d'abord, après avoir acquis l'aspect germinal, apparaissent les spermatogonies qui émergent de l'épithélium germinatif pour occuper la zone médullaire des tubules gonadiques et les cellules basales pariétales servant de réservoirs trophiques (cellules nourricières) aux spermatogonies.
2. Après que la multiplication spermatogoniale ait comblé la lumière tubulaire, se différencient au sein de l'épithélium germinatif des protogonies qui s'accroissent en ovocytes primaires s'entourant de cellules folliculeuses tout en restant cantonnées sur la paroi (voir schéma ci-dessous).
En conclusion :
L'analyse du cycle sexuel de l'Escargot, Helix aspersa, appuyée sur l'observation histologique et ultrastructurale de la gonade a permis de définir les modalités de l'expression de l'hermaphrodisme au niveau de l'ovotestis adulte ou postembryonnaire.
1. Les cellules qui composent l'épithélium germinatif, foyer de la gamétogenèse sont-elles sexuellement indifférenciées ? Différentes constatations appuient cette hypothèse :
- Dans l'ovotestis, il n'y a pas de régions spécialisées dans la production des gamètes de chaque sexe.
- Les cellules de l'épithélium germinatif sont morphologiquement identiques.
- Les deux catégories de gamètes ainsi que leurs cellules accessoires proviennent de cellules souches semblables à des cellules somatiques.
D'après ces résultats et dans les limites de l'observation histologique et de l'examen ultrastructural, il semble que la cellule souche soit réellement une cellule pluripotente pouvant engendrer indifféremment une cellule germinale ou une cellule somatique.
Cette constatation met à mal la théorie de Weismann qui considère qu'il existe une ségrégation entre tissu germinal (ou germen) qui va engendrer les gamètes et les tissus qui vont engendrer le reste du corps (ou soma). On sait maintenant que cette distinction n'est plus valable puisqu'on peut cloner des cellules somatiques totipotentes qui seront capables d'engendrer n'importe quelle catégorie cellulaire, y compris la lignée germinale.
2. En ce qui concerne l'observation des mécanismes de la sexualisation, l'orientation dans le sens mâle se caractérise par une phase germinative qui aboutit à une multiplication spermatogoniale envahissant la cavité gonadique.
L'ovogenèse qui succède toujours à la spermatogenèse démarre à partir de cellules indifférenciées. Chez Helix aspersa, l'ovogenèse se distingue par l'absence de toute division cellulaire.
3. L'expression de la sexualité au sein de la gonade ainsi que la cytodifférenciation des gamètes semblent répondre à un certain nombre de stimuli dont l'origine ne peut être saisie sur le simple plan de l'observation. La cellule germinale paraît sexuellement indifférenciée. Mais, pour concevoir le processus de sa sexualisation, il est nécessaire de faire appel à l'action d'agents humoraux extérieurs à la cellule de l'épithélium germinatif. Le sexe de la cellule germinale semble conditionné par le contexte tissulaire et humoral qui ne serait pas identique au moment de l'apparition de chacune des lignées sexuelles.
Pour étayer cette hypothèse, il est nécessaire d'apporter des arguments d'ordre physiologique.
Il faudra d'abord tester les potentialités propres de la gonade en l'isolant de son contexte tissulaire et humoral. Pour ce faire, on va employer une technique extrêmement performante. Il s'agit de la culture organotypique. Il faudra observer les réactions de l'épithélium germinatif livré à lui-même sur un milieu de culture anhormonal.
Ensuite, afin de mettre en évidence l'influence de facteurs humoraux sur la gonade, on va soumettre la gonade indifférenciée à l'influence d'organes potentiellement actifs en les associant directement la gonade et ces organes en culture. Cet aspect du problème fera l'objet d'un prochain article qui aboutira à des résultats surprenants.
Sources :
- Guyard A. (1971) - Etude de la différenciation de l'ovotestis et des facteurs contrôlant l'orientation sexuelle des gonocytes de l'Escargot Helix aspersa Müller. Thèse d'État soutenue à la Faculté des Sciences de l'Université de Franche-Comté.
- Documents inédits tirés de la photothèque personnelle de André Guyard.








 Facebook | | |
Facebook | | |
Les commentaires sont fermés.