



04/11/2009
L'Escargot (15) Conclusion générale
 L'Escargot (15) : Interprétation de l'hermaphrodisme chez Helix aspersa
L'Escargot (15) : Interprétation de l'hermaphrodisme chez Helix aspersa
par André Guyard
(suite de l'Escargot 14)
Le présent article constitue une conclusion aux 14 articles précédents consacrés à l'Escargot et au déterminisme du sexe chez cet animal hermaphrodite.
Le comportement de la gonade d'Escargot en culture organotypique autorise à penser que la différenciation sexuelle des gonocytes est sous la dépendance d'une séquence hormonale. La confrontation des résultats de l'analyse du cycle in vivo et de l'étude expérimentale in vitro permet de mieux comprendre certaines modalités de l'expression de l'hermaphrodisme.
1) La culture de l'ébauche gonadique isolée démontre que non seulement chaque cellule-souche morphologiquement identique à une cellule somatique est sexuellement bivalente mais qu'elle est potentiellement une cellule femelle. Chez cette espèce hermaphrodite, le sexe femelle se réalise dans des conditions anhormonales. Il s'agit d'une autodifférenciation femelle.
2) Comment interpréter la protandrie ? Il faut supposer que la phase germinative qui débouche in vivo sur la spermatogenèse est déclenchée par un facteur masculinisant. L'expérimentation nous apprend que ce facteur présumé est humoral. Il est présent dans l'hémolymphe d'animaux en phase mâle et il permet l'accomplissement de la spermatogenèse in vitro. La sécrétion de cet agent est discontinue puisque sa présence dans l'hémolymphe est épisodique.
Le primum movens de l'action androgène siège au niveau du ganglion cérébroïde d'animaux d'âge postembryonnaire puisque cet organe favorise la multiplication spermatogoniale et, conjointement avec le tentacule oculaire, empêche la manifestation de l'autodifférenciation ovocytaire. Un fonctionnement endocrine discontinu du cerveau expliquerait les démarrages de spermatogenèses abortives au cours du premier mois de la vie postembryonnaire.
3) L'apparition des ovocytes un mois après l'éclosion correspondrait à un épuisement corrélatif des hormones masculinisantes cérébrales. La différenciation des protogonies dans le sens femelle coïncide dans un tubule donné avec l'arrêt de la multiplication spermatogoniale. In vitro, c'est l'époque où la seule présence des ganglions cérébroïdes ne permet plus d'assumer le contrôle de la spermatogenèse et de refréner l'autodifférenciation ovocytaire. Le taux de facteur masculinisant serait insuffisant pour imposer le sexe mâle à toute la gonade puisqu'il s'établit une espèce de gradient de sexualité dans la culture. La partie la plus proche du tissu nerveux est occupée par des spermatogonies en multiplication alors que dans la partie la plus éloignée, des gonocytes échappant probablement à l'action du facteur masculinisant s'autodifférencient en ovocytes.
4) Plus tard, au cours du cycle annuel de la gonade mature, le démarrage de la spermatogenèse puis l'apparition des ovocytes sur les parois peuvent s'expliquer de la même façon par l'intervention puis l'épuisement du facteur androgène. Or, chez les Crustacés, le cerveau qui contrôle le fonctionnement de la glande androgène a lui-même le même effet directement sur les gonocytes (J. Berreur-Bonnenfant, 1968). L'association in vitro des ganglions cérébroïdes d'animaux en spermatogenèse avec la gonade ne permet jamais la réalisation d'une spermatogenèse comparable à celle réalisée par l'adjonction d'hémolymphe d'hiver. Peut-être que cette expérience ne met en évidence que la stimuline contrôlant le fonctionnement d'un hypothétique tissu androgène.
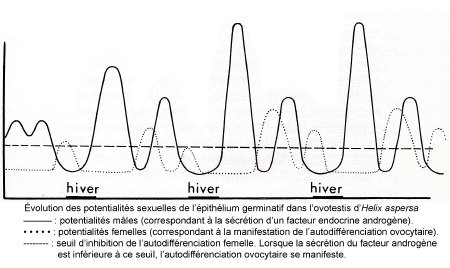
5) En outre, après accomplissement de la spermatogenèse, la lignée femelle poursuit son évolution. Or in vitro, le cerveau extirpé d'animaux en ovogenèse ne permet que la seule manifestation de la lignée femelle. C'est une nouvelle preuve de la labilité du facteur cérébral masculinisant. En sa présence en un taux suffisant, la lignée mâle entre en multiplications spermatogoniale ; en son absence ou en sa présence en quantité insuffisante, la lignée femelle se manifeste et amorce son auxocytose.
6) On peut se demander quel est le mécanisme responsable du déclenchement et de la modulation de la sécrétion de ce facteur masculinisant. D'une façon générale, il est certain que le système nerveux intervenant aussi bien dans le contrôle de la vie de nutrition et de reproduction que dans la vie de relation de l'organisme, la sécrétion des neurohormones est sousmise à l'influence des facteurs exogènes trophiques, climatiques ou psychiques, les récepteurs sensoriels (notamment le tentacule oculaire des Gastéropodes) pouvant jouer le rôle de relais. Ces considérations expliqueraient l'intervention des facteurs externes dans l'expression du phénotype cytosexuel chez les espèces hermaphrodites.
On peut également envisager que le signal du virage sexuel au niveau du cerveau provienne de l'ovotestis lui-même qui, par une sorte de feed-back déclenché par exemple par l'état de réplétion des tubules gonadiques dû à l'abondance des spermatogonies, avertisse le système nerveux central par voie humorale.
7) La participation d'autres facteurs humoraux contrôlant la physiologie de l'une ou l'autre des gamétogenèses doit être supposée pour expliquer la coexistence des deux lignées gamétiques.
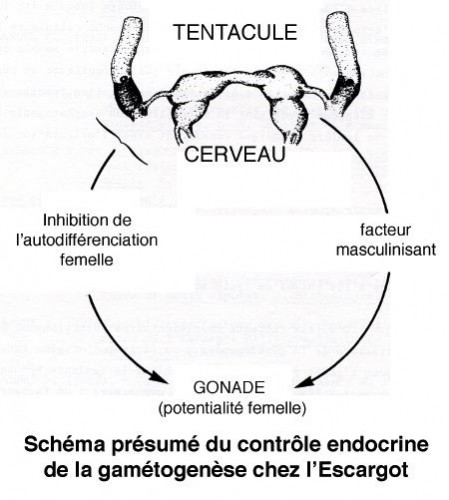
Ainsi, pendant que se déroule la spermiogenèse in vivo ou en culture en présence d'hémolymphe d'hiver, l'accroissement ovocytaire est réprimé. C'est alors qu'on peut supposer l'intervention d'une inhibition de l'ovogenèse parallèlement à l'action d'un facteur spermiogénétique d'origine inconnue. Le fait que le tentacule montre un rôle constamment répressif sur l'ovogenèse est un indice en faveur de l'intervention de cet organe dans l'inhibition de l'accroissement ovocytaire.
D'autre part, un facteur cérébral semble, contrôler l'auxocytose. En culture, le ganglion cérébroïde prélevé sur des animaux en ovogenèse a une influence bénéfique sur l'accroissement ovocytaire sans qu'on puisse préjuger d'un rôle sur la vitellogenèse.
On peut soupçonner l'intervention de multiples facteurs dont les interactions constituent un jeu subtil permettant l'évolution en parallèle des deux lignées sexuelles ; ainsi la préméiose au sein des ovocytes débute fréquemment en même temps que s'amorcent les divisions réductionnelles des spermatocytes. Peut-être y a-t-il là une indication de l'intervention d'un facteur méiotique non sexualisé ?
Le schéma de la sexualisation des cellules germinales au sein de la gonade paraît plus simple. Les protogonies ou gonies indifférenciées sont sexuellement bivalentes. Elles reçoivent de la part du soma l'impulsion qui les orientera vers un sexe ou vers l'autre. C'est la nature et le cheminement de cette impulsion qui sont en cause.
Chez Helix aspersa, la culture organotypique a permis de mettre en évidence le phénomène d'autodifférenciation femelle.
Le schéma de la séquence hormonale qui contrôle la différenciation dans le sens mâle, évoqué par les résultats de l'association de la gonade avec le ganglion cérébroïde demande à être confirmé. S'agit-il de facteurs qualitativement distincts ou simplement de la modulation d'un unique facteur stimulateur de la gamétogenèse ?
Il ne faut pas perdre de vue que ces résultats ont été obtenus avec la seule technique des cultures organotypiques. La mise en évidence d'actions endocrines ne pourra être résolue qu'avec le concours d'expériences complémentaires. Injections d'extrait cérébral et/ou tentaculaire, ablation sélective de cellules nerveuses, greffes ou implantations de ganglions, observation de la différenciation fine au niveau des organites des cellules germinales, identification chimique des substances actives constituent autant de techniques à exploiter pour poursuivre la discussion sur le contrôle de l'hermaphrodisme chez l'Escargot.
RÉSUMÉ
Helix aspersa Müll. est une espèce d'Hélicidés hermaphrodite protandre. L'étude histologique et ultrastructurale complétée par la mise en culture organotypique de la gonade a permis d'apporter quelques précisions quant aux mécanismes qui régissent la sexualisation des gonocytes.
La mise en place de la gonade est caractérisée par une période d'indifférenciation qui se prolonge jusqu'à dix jours après l'éclosion. Il n'existe qu'une seule souche cellulaire à l'origine des deux lignées germinales.
La protandrie semble due à une induction mâle d'origine cérébrale qui se manifeste en réprimant une tendance spontanée à l'autodifférenciation femelle et par un effet mitogène favorisant la multiplication spermatogoniale qui envahit la lumière gonadique consacrant ainsi l'orientation définitive du gonocyte dans le sens mâle.
Un mois après l'éclosion, l'hermaphrodisme se manifeste par la différenciation d'ovocytes en position pariétale. Ce phénomène réalité de l'autodifférenciation ovocytaire non refrénée par le cerveau dont l'action modulée selon l'âge ou la saison déterminera les phases du cycle sexuel.
D'autres facteurs complémentaires interviennent dans la réalisation de la gamétogenèse : un facteur d'origine tentaculaire dont l'effet principal est d'inhiber la tendance spontanée des gonocytes à l'autodifférenciation ovocytaire ; un facteur dont le siège est le ganglion cérébroïde qui favorise l'accroissement de la cellule femelle avec le concours de facteurs trophiques.
09:59 Publié dans Biologie, Invertébrés, Mollusques, Sexualité-embryologie | Tags : escargot, sexualité, hermaphrodisme | Lien permanent | Commentaires (0) |  Facebook | | |
Facebook | | |
Les commentaires sont fermés.