



13/11/2009
L'Escargot (6) : L'hermaphrodisme
Étude en microscopie photonique et électronique de la glande hermaphrodite de l'Escargot : les gamètes mâles et femelles sont produits dans chaque tubule de la gonade où l'on trouve simultanément les ovocytes sur la paroi et les gonocytes mâles en évolution dans la lumière des tubules.
 L'Escargot (6) :
L'Escargot (6) :
L'hermaphrodisme
Généralités
Par André Guyard
(suite de l'Escargot 5)
Animal hermaphrodite, l'Escargot constitue un modèle incomparable pour l'étude de la sexualité. Cet hermaphrodisme appelle une foule de questions.
Depuis que le microscope existe, les chercheurs ont tenté d'observer les cellules de la "génération" et la sexualité constitue l'un des chapitres les plus attractifs de la biologie contemporaine.
Le plus souvent, chez les espèces à sexes séparés (espèces gonochoriques), de nombreux caractères distinguent l'aspect mâle de l'aspect femelle et le biologiste sait que cette discrimination s'applique jusque dans l'intimité de chaque cellule - en particulier au niveau des chromosomes sexuels - et que la détermination est très précoce : dès la fécondation de 1'ovule par le spermatozoïde, le sexe est établi.
Cependant, de nombreux organismes présentent un mélange des caractères des deux sexes et produisent les deux catégories de gamètes. Les formes hermaphrodites où les modalités de la sexualité revêtent de nombreux aspects sont particulièrement favorables à l'analyse des mécanismes de la sexualisation.
Définition de l'hermaphrodisme chez les Mollusques Gastéropodes
L'hermaphrodisme est une caractéristique générale des Gastéropodes pulmonés. Les modalités de la sexualité revêtent tant d'aspects chez les formes hermaphrodites que les auteurs se sont efforcés d'établir un classement des différents types rencontrés.
P. Pelseneer (1894) considère comme hermaphrodites les Mollusques qui présentent en même temps les gamètes mâles et femelles. Pour W. R. Coe (1945), l'hermaphrodisme est un phénomène essentiellement physiologique. Selon Coe, quatre éventualités peuvent se présenter chez les Mollusques hermaphrodites :
1) l'hermaphrodisme fonctionnel où gamètes mâles et femelles mûrissent au cours de la même période ;
2) la sexualité consécutive avec un seul virage sexuel séparant deux périodes de vie sexuellement différentes ;
3) la sexualité rythmique consécutive caractérisée par des phases sexuelles différentes au cours de périodes de reproduction successives ;
4) la sexualité alternative avec des phases soit mâles soit femelles se succédant dans n'importe quel ordre au cours de périodes de reproduction successives.
Déterminer un type de sexualité selon cette classification implique des études de populations. Cette étape est indispensable pour les espèces "faussement gonochoriques" qui rentrent dans la catégorie 2 de COE. C'est le cas de nombreux Prosobranches dont Crepidula (W. R. Coe, 1938, 1942), Calyptraea sinensis L. (G. Bacci, 1951 ; W. Streiff, 1967) ou Patella vulgata L. (J. H. Orton, 1920, 1946 ; G. Bacci, 1952 ; M. Choquet, 1966, 1969).
Ainsi, la notion d'hermaphrodisme n'est pas facile à définir. Il faut admettre comme hermaphrodite tout individu qui présente des cellules germinales de chacun des deux sexes mûrissant dans une même période ou dans des périodes successives de son cycle vital. Cette définition englobe aussi bien les animaux dont les produits génitaux mûrissent ensemble que les "faux gonochoriques" apparemment unisexués.
Afin de regrouper dans une série naturelle tous les cas d'hermaphrodisme inclus dans cette définition, G. Bacci (1951) propose la distinction entre hermaphrodites simultanés et hermaphrodites successifs.
Les hermaphrodites simultanés sont caractérisés par la maturation synchrone de leurs deux sortes de gamètes.
Parmi les hermaphrodites successifs, G. Bacci regroupe :
- les cas de maturation asynchrone où gonocytes mâles et femelles coexistent mais ne mûrissent pas simultanément et
- les cas de maturation distincte où phase mâle et phase femelle sont bien séparées dans le temps.
Cas de l'Escargot
Montrant un cas d'hermaphrodisme à maturation asynchrone des gonocytes mâles et femelles, l'Escargot constitue un modèle de choix permettant d'étudier comment s'effectuent l'orientation des gamètes et le phénomène de sexualisation. En effet, l'arbre génital de l'escargot tout entier est marqué par l'hermaphrodisme. Ce gastéropode est muni d'un ovotestis, c'est-à-dire que dans sa gonade mûrissent les deux lignées de gonocytes (cellules sexuelles). L'ovotestis est réellement une glande hermaphrodite qui produit à la fois des spermatozoïdes et des ovules.
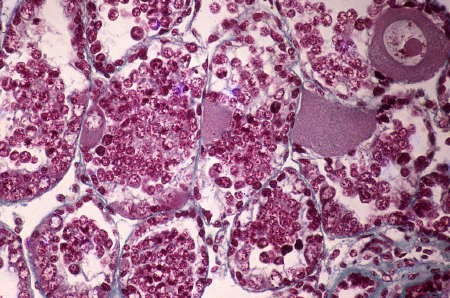
Cette glande hermaphrodite montre une structure étonnante : elle est formée d'une série de tubules qui tous renferment à la fois les cellules sexuelles : gonocytes mâles et gonocytes femelles, c'est-à-dire les gamètes en maturation qui vont aboutir à l'élaboration des spermatozoïdes, d'une part et des ovules, d'autre part.
Dans le cas de l'Escargot, une série de questions se pose :
1. Comment se fait-il qu'il n'y ait pas fécondation interne puisque les gonocytes se côtoient dans les mêmes acini ? Cette question ne se pose pas pour de nombreux organismes animaux hermaphrodites où l'appareil génital est composé d'une part de testicules qui vont fournir des spermatozoïdes, et d'autre part d'ovaires qui produisent des ovules. C'est le cas, par exemple des lombrics ou des sangsues.
2. Les gonocytes mâles proviennent-ils de lignées cellulaires différentes ou bien de cellules souches identiques ? Dans ce cas, quels sont les phénomènes qui vont prévaloir dans l'orientation sexuelle des gonocytes ?
Cette question est fondamentale en biologie. Au cours de l'embryogenèse, le biologiste allemand F. L. A. Weismann a émis en 1883 l’hypothèse de l’existence d’un plasma germinatif immortel ou germen à l'origine de la gamétogenèse qui passerait de génération en génération et serait opposé aux cellules du restant de l'organisme ou soma, qui correspondrait à la partie mortelle de l'individu.
P. Laviolette (1954a) aborde la question par le côté expérimental. Grâce à des techniques chirurgicales minutieuses, ce dernier auteur montre que le tissu gonadique des Arionidés peut régénérer après castration à partir de cellules déjà engagées dans la voie somatique ; résultat important qui oblige à reconsidérer la théorie de la lignée germinale de Weismann.
L'avènement du microscope électronique permet à P.-P. Grassé, N. Carasso et P. Favard (1956) d'explorer les ultrastructures cellulaires au cours de la spermiogenèse de l'Escargot Helix aspersa Mull.
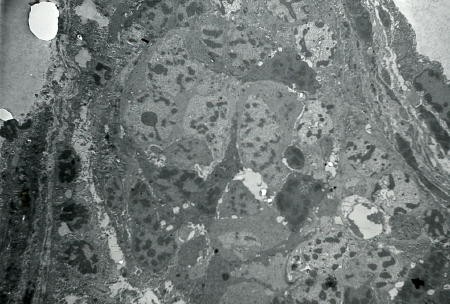
Parallèlement aux travaux menés chez les Vertébrés, l'idée d'un contrôle endocrine de la sexualité fait son apparition. R. Aubry (1962), par des injections d'hormones sexuelles stéroïdes chez Limnea stagnalis L. constate des modifications au niveau des gamètes.
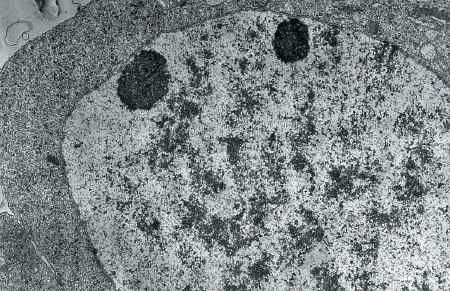
Par des expériences d'ablation tentaculaire et d'injection d'extraits cérébral et tentaculaire, D. Pelluet et N. J. Lane (1961) éprouvent l'influence endocrine du cerveau et du tentacule sur la gamétogenèse de limaces. Ils soupçonnent l'existence de deux hormones antagonistes, l'une d'origine cérébrale favorisant l'ovogenèse, l'autre d'origine tentaculaire inhibant la lignée femelle.
Avec la culture organotypique vient une abondance de travaux qui d'abord s'intéressent à l'embryologie des Vertébrés puis s'étendent à l'endocrinologie des Invertébrés. En conjuguant les observations aux microscopes photonique et électronique et les techniques de culture organotypique, j'ai entrepris l'étude de la glande hermaphrodite d'Helix aspersa Müller, l'Escargot Petit-Gris.
Il importait tout d'abord de définir l'état hermaphrodite de la gonade et de déterminer le mécanisme par lequel une cellule germinale s'oriente vers un sexe ou vers l'autre. L'étude histologique et ultrastructurale du cycle de la gonade constitue l'objet des deux prochains chapitres.
Grâce à la culture de la gonade isolée ou au contact d'organes présumés endocrines, j'ai tenté de déceler les mécanismes qui contrôlent l'expression de l'hermaphrodisme chez l'Escargot. Ce sera l'objet de chapitres ultérieurs.
La suite l'Escargot 7.
- Documents inédits tirés de la photothèque personnelle de André Guyard.
14:56 Publié dans Biologie, Invertébrés, Mollusques, Sexualité-embryologie | Tags : sexualité, hermaphrodisme, escargot, mollusques, gamétogenèse | Lien permanent | Commentaires (0) |  Facebook | | |
Facebook | | |
Les commentaires sont fermés.