
L'escargot (5) : Biologie et développement
14/11/2009
Bien qu'ils soient hermaphrodites et et que leur gonade produise les deux sexes gamétiques, il n'y a pas autofécondation chez les escargots. L'accouplement est réciproque avec échanges de spermatophores mis en réserve chez le partenaire, en attente de la maturation des ovocytes.
 L'Escargot (5) : Biologie et développement
L'Escargot (5) : Biologie et développement
par André Guyard
(suite de l'Escargot 4)
L'Escargot, mollusque terrestre, respire l'air en nature par sa cavité palléale dont le toit présente des mouvements rythmiques ; ces mouvements ne sont actifs que par temps humide ; la dessication du milieu détermine le passage à la vie ralentie ; l'animal est, de même que pendant l'hiver, complètement rétracté à l'intérieur de sa coquille ; l'épiphragme peut fermer l'ouverture de celle-ci.
Les déplacements se font par reptation sur le pied, facilitée par le mucus ; la longueur du pied ne varie pas, mais sa surface est parcourue par des ondes de contraction des muscles qui se propagent d'avant en arrière (voir la locomotion chez l'Escargot, chapitre III).
L'animal présente un phototactisme variable suivant les conditions physiologiques.
L'alimentation est faite de tissus végétaux tendres, feuilles surtout.
À la mauvaise saison, l'Escargot de Bourgogne (Helix pomatia) sécrète un épiphragme, une sorte d'opercule calcaire qui ferme le péristome et s'enterre. En revanche, le Petit-Gris (Helix aspersa) hiberne en recherchant un refuge dans des pierriers et en se collant au support.
C'est au Petit-Gris que nous nous intéresserons plus spécialement car son élevage est relativement aisé.

La durée de l'hiver comtois abrège son cycle annuel. Le réveil du Petit-Gris a lieu fin mars-début avril. Le processus de réveil est certainement déclenché par les conditions climatiques : hausse des températures, allongement de la durée du jour… avec peut-être un relais vers les ganglions cérébroïdes par les ganglions oculaires.
Bien qu'hermaphrodites, les escargots s'accouplent. Sous nos latitudes, la période des accouplements se situe au mois de juin.
L'hermaphrodisme est protandre et l'accouplement a lieu lorsque seuls les gonocytes mâles sont mûrs. Les spermatozoïdes sont agglutinés en spermatophores par la glande du fouet.
Il est probable que la gamétogenèse avec protandrie est initiée par des processus neurosécrétoires des ganglions cérébroïdes comme le montrent les expériences que j'ai réalisées en culture organotypique en associant gonade indifférenciée et collier périœsophagien. Dans d'autres expériences in vitro, le mésocérébron droit a été proposé comme centre de contrôle du comportement sexuel.

L'accouplement est réciproque et chaque animal excite l'autre en lui piquant la peau avec son dard, qui se brise et sort par l'orifice génital grâce au mucus des glandes multifides.
Dans sa thèse (1999)[1], Joris M. Koene analyse l'emploi du dard, qui constitue une composante insolite du comportement sexuel de certaines espèces d'Hélicidés dont Helix aspersa, qui enfonce cet organe composé de calcium et recouvert de mucus dans la peau de son partenaire au moment de l'accouplement.
Cette thèse traite des raisons de ce comportement et de la manière dont le système nerveux central le contrôle. Les résultats démontrent que l'action du dard ne constitue pas seulement un simple apport de calcium parce qu'il est rarement incorporé par le récipiendaire et parce qu'il ne contient qu'une petite quantité de calcium. Cependant, la plupart des dards qui sont utilisés perforent la peau et entrent en contact avec l'hémolymphe. Le mucus qui est produit par les glandes digitiformes et recouvre le dard contient une substance qui affecte les organes reproducteurs. L'effet observé suggère que, lorsque le dard est reçu, davantage de sperme est emmagasiné par le récepteur. Cet effet est important pour la compétition du sperme parce que les escargots entreposent le sperme de plusieurs partenaires avant de pondre leurs œufs, L'introduction d'une substance bioactive par le dard peut être expliquée soit comme un choix de partenaire ou comme une manipulation du partenaire.
L'auteur conclut que l'escargot se sert de son dard pour transférer une substance dans le partenaire, afin de moduler l'emmagasinage du sperme donné. Il faut noter que certaines espèces transfèrent des substances bioactives à leurs congénères. Il propose d'utiliser le terme "allohormone" pour désigner ces substances, les distinguant des hormones et phéromones.
Pour analyser le rôle du cerveau dans l'emploi du dard et les autres composantes de l'accouplement, il a utilisé une approche in vivo. Il démontre, en procédant à des stimulations et enregistrements électriques de neurones du mésocérébron droit dans l'animal intact, que ces neurones sont impliqués dans le lancement du dard et l'éversion du pénis. Il teste aussi l'hypothèse que certains neuropeptides sont impliqués dans différents aspects du comportement sexuel. Il démontre que l'APGWamide est responsable de l'éversion génitale. Ces résultats, jumelés aux résultats disponibles et obtenus dans d'autres mollusques, concluent que la région antéromédiale du ganglion cérébral droit est une région du cerveau qui a été conservée dans l'évolution pour contrôler le comportement sexuel.
[1] Joris M. Koene (1999) - Behavioural and neurobiological aspects of dart shooting in the garden snail Helix aspersa, Ph.D. Thesis McGill University.
Puis le pénis est dévaginé en doigt de gant (éversion) et introduit dans le vagin de l'autre escargot : il y dépose les spermatophores qui sont emmagasinés dans le réceptacle séminal jusqu'à la maturation des ovules. La fécondation de ceux-ci a lieu dans le vagin.
Les œufs, très riches en vitellus, sont des perles sphériques de 1 mm de diamètre, pondus au nombre d'une centaine dans un trou creusé par l'animal. Ils sont entourés d'une coque résistante. Les pontes de Petits-Gris sont commercialisées sous le nom de caviar d'escargot.
La ponte survient un à deux mois après, soit en juillet-août. Le Petit-Gris dépose dans son trou 120 à 160 œufs de 4 mm de diamètre. La durée de l'incubation dépend de la température, l'éclosion survenant trois semaines à un mois plus tard et s'étalant sur plusieurs jours. Pris au nid, les jeunes mesurent de 3,5 à 4 mm de plus grand diamètre.


Le développement est condensé par rapport à celui des Gastéropodes marins ; l'éclosion de l'œuf donne directement un jeune Escargot. L'Escargot étant un animal terrestre, son développement ne présente pas de larve trochophore.

Pendant une dizaine de jours, sans se nourrir, ils résorbent leur poche vitelline à laquelle se substitue progressivement la glande digestive ou hépatopancréas. Ce stade se traduit macroscopiquement par la pigmentation du tortillon qui, de blanc albumineux, devient jaune puis marron.
L'hépatopancréas étant constitué, l'animal peut alors se nourrir. Le jeune entre en hibernation tardivement. Alors que l'adulte s'enfouit en octobre, il n'est pas rare de rencontrer des jeunes de 5 à 6 mm jusque vers la mi-novembre.
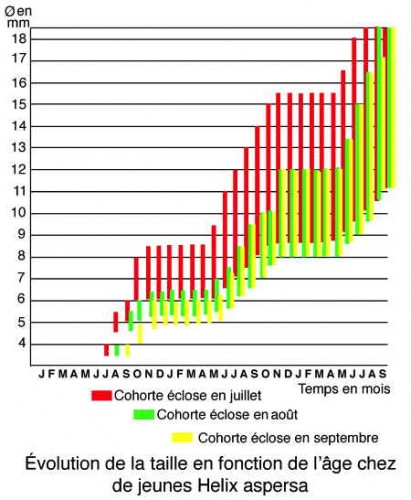
Chez le Petit-Gris, le développement se poursuit jusqu'à la mauvaise saison où la courbe taille-poids cesse d'évoluer. Puis le développement reprend au printemps. Mais une sécheresse peut provoquer une estivation avec un arrêt de la croissance. En deux ans, le Petit-Gris atteint une taille de 20 mm (voir courbes de croissance de trois cohortes successives sur le diagramme ci-dessous).
Voir la suite l'Escargot 6.
- Documents inédits tirés de la photothèque personnelle de André Guyard.
Les commentaires sont fermés.