
L'escargot (4) : Anatomie
15/11/2009
Les Gastéropodes sont vraiment des animaux "tordus". Au cours de leur embryogenèse, ils subissent flexion et torsion, voire enroulement dans une coquille spiralée… Et en plus, un appareil génital à tout faire : spermatozoïdes et ovules.
 L'Escargot de Bourgogne (4)
L'Escargot de Bourgogne (4)
Helix pomatia
Anatomie interne
par André Guyard
(suite de l'Escargot 3)
Avant d'aborder l'anatomie proprement dite de l'Escargot de Bourgogne, il convient de donner quelques explications sur le développement des Gastéropodes.
Généralités sur le développement des Gastéropodes
Cette classe rassemble des Mollusques à morphologie externe assez uniforme, mais assez différents par leur anatomie interne. Ils présentent un pied aplati en forme de sole servant à la reptation ; la tête, bien développée, montre la bouche munie d'une radula, sorte de langue munie de dents et les tentacules sensoriels. La masse viscérale, enroulée en hélice, est généralement protégée par une coquille univalve également hélicoïdale, mais plusieurs groupes marquent une tendance à la réduction de cette coquille.
L'anatomie interne montre une dissymétrie tout à fait remarquable qui résulte de modifications subies par la masse viscérale au cours du développement, et qu'on peut schématiser ainsi :
1° Tout d'abord, une flexion du tube digestif, par développement de la masse viscérale en hauteur dans le sens dorso-ventral ; le tube digestif prend la forme d'un U, ses deux extrémités, antérieure et postérieure, bouche et anus, étant assez rapprochées.
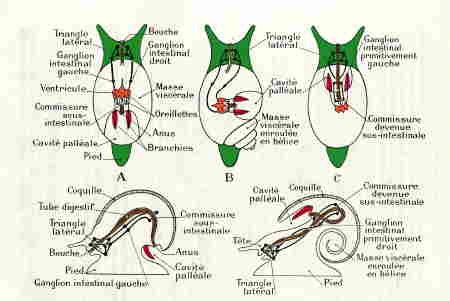
2° Puis, une torsion de la masse viscérale par rapport à la région céphalo-pédieuse ; cette torsion, de 180° autour de son axe, s'effectue en sens inverse des aiguilles d'une montre pour un observateur regardant l'animal par sa face dorsale et vers l'avant ; elle ramène en avant l'ouverture de la coquille et la cavité palléale avec ses dépendances (complexe palléal), situées primitivement en arrière. L'anus est ainsi rapproché de la bouche, et le tube digestif subit cette torsion. Les organes pairs (oreillettes, branchies, rein), situés primitivement à gauche, passent à droite et réciproquement.
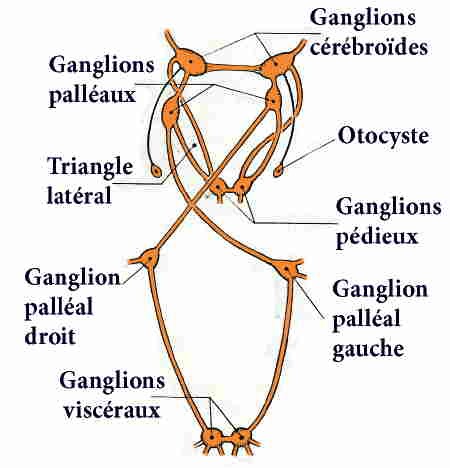
Le système nerveux présente également les traces de cette torsion : la région antérieure, céphalique, avec les triangles latéraux, n'en est pas affectée, mais la chaîne nerveuse ventrale la subit ; le ganglion intestinal primitivement droit est amené à gauche en passant au-dessus du tube digestif et le ganglion intestinal primitivement gauche est amené à droite en passant au-dessous du tube digestif; la commissure primitivement sous-intestinale passe au-dessus du rectum; la chaîne nerveuse ventrale est alors croisée en X ou en 8, le connectif partant du ganglion palléal gauche passant sous le tube digestif, et celui partant du ganglion palléal droit passant au-dessus.
L'étude du développement montre que cette torsion est un phénomène brutal, ne durant que quelques minutes, qui affecte la larve véligère chez les Gastéropodes marins.
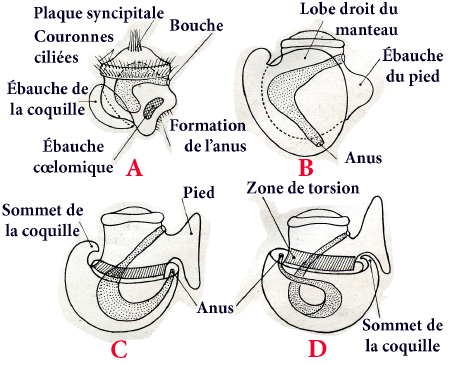
3° Enfin la masse viscérale s'enroule en hélice sur le côté devenu droit après torsion ; cet enroulement en hélice s'effectue autour d'un axe oblique par rapport à l'axe de torsion ; il détermine l'atrophie du côté adjacent à cet axe, c'est-à-dire le côté devenu droit après la torsion ; les organes pairs de ce côté, comme l'oreillette, la branchie, le rein (primitivement gauches), sont atrophiés et disparaissent plus ou moins. Cet enroulement est tout à fait indépendant de la torsion, qu'il précède d'ailleurs.
Ces modifications sont plus ou moins accentuées et nettes suivant les divers groupes de Gastéropodes.
On peut admettre que les Gastéropodes actuels dérivent d'animaux symétriques, formant le groupe des Isopleures (à flancs égaux), actuellement fossiles comme les Bellerophon. L'anatomie interne de ces animaux est inconnue, mais leur coquille parfaitement symétrique est enroulée dans le plan de symétrie, en spirale (Planspiralia), par opposition aux Turbospiralia, enroulés en hélice. L'anatomie interne des Gastéropodes actuels ou Anisopleures (à côtés inégaux) varie selon la sous-classe :
1° Les Prosobranches
caractérisés par la disposition antérieure de la cavité palléale, les branchies étant situées en avant du cœur, d'où leur nom. Leur chaîne nerveuse montre le croisement en X caractéristique, et on les appelle parfois Streptoneures. Ce sont les Gastéropodes les plus primitifs. Les sexes sont séparés. On les oppose aux deux autres sous-classes, réunies parfois sous le nom d'Euthyneures, car leur chaîne nerveuse n'est pas croisée, et dans lesquelles les espèces sont hermaphrodites.
2° Les Opisthobranches
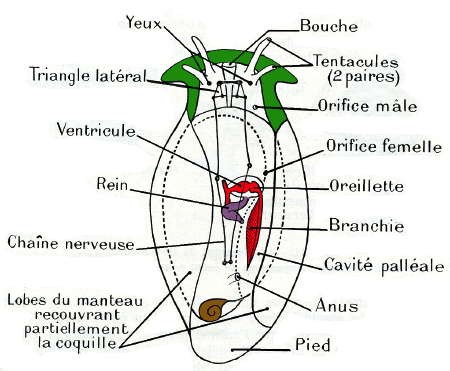
Ces Gastéropodes tous marins n'ont qu'une oreillette, en arrière du ventricule, et qu'une branchie, située en arrière du cœur, d'où leur nom. La cavité palléale, largement ouverte, est située sur le côté droit et en arrière. La masse viscérale et la coquille sont souvent très réduites, et l'apparence extérieure est souvent à peu près symétrique. Mais l'absence d'un rein, d'une oreillette et d'une branchie, montre que les Opisthobranches dérivent de formes ayant subi l'enroulement et la torsion, comme les Prosobranches ; à partir de cette disposition primitive, ils ont subi un phénomène secondaire de détorsion, dans le sens des aiguilles d'une montre, qui a ramené la cavité palléale en arrière du corps, la branchie en arrière du cœur et l'oreillette en arrière du ventricule : la chaîne nerveuse n'est plus croisée. On peut suivre cette évolution secondaire sur une série de formes, et en particulier au niveau de la chaîne nerveuse qui présente toujours une dissymétrie.
3° Les Pulmonés
Ces Gastéropodes ont perdu leurs branchies et leur cavité palléale est transformée en un poumon : formes d'eau douce ou terrestres, ils respirent l'air en nature. C'est dans cette sous-classe que l'on rencontre nos escargots.
Les Pulmonés ont subi la torsion, car la cavité palléale est placée en avant ; mais leur système nerveux est concentré dans la tête (processus de céphalisation), et la chaîne nerveuse ventrale presque inexistante a échappé à la torsion. Il n'y a qu'un rein et qu'une oreillette placée en avant du ventricule.
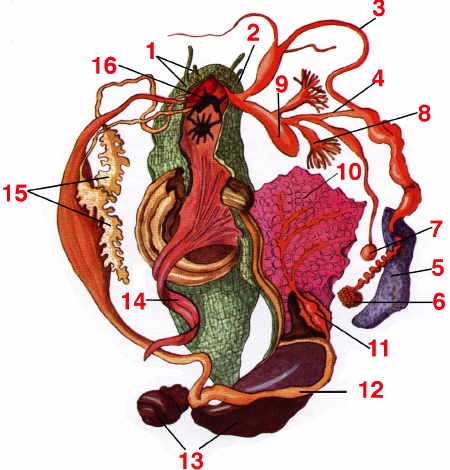
Dissection de l'Escargot de Bourgogne
1 : tentacules ; 2 : orifice génital ; 3 : canal déférent ; 5 : glande de l'albumine ; 6 : glande hermaphrodite ; 7 : spermathèque ; 8 : glande multifide ; 9 : poche du dard ; 10 : poumon ; 11 : cœur ; 12 : appareil digestif ; 13 : hépato-pancréas ; 14 : muscle columellaire ; 15 : glandes salivaires ; 16 : ganglions cérébroïdes
Anatomie de l'Escargot
Téguments
Ils sont formés par l'épiderme, simple, caractérisé par l'abondance des glandes à mucus, et un derme à muscles lisses bien développés, surtout pour former les muscles de la reptation et le muscle columellaire ; celui-ci s'attache d'une part sur la columelle et, d'autre part, s'irradie dans la tête et le pied, permettant leur rétraction à l'intérieur de la coquille.
Système nerveux
Chez l'Escargot et les Gastéropodes pulmonés, le système nerveux est caractérisé par une très forte condensation céphalique : tous les ganglions se trouvent localisés dans la région du bulbe pharyngien, formant une masse qui entoure l'œsophage. La région dorsale de cette masse correspond aux ganglions cérébroïdes réunis par une commissure très courte et innervant les organes de la tête et le pénis.
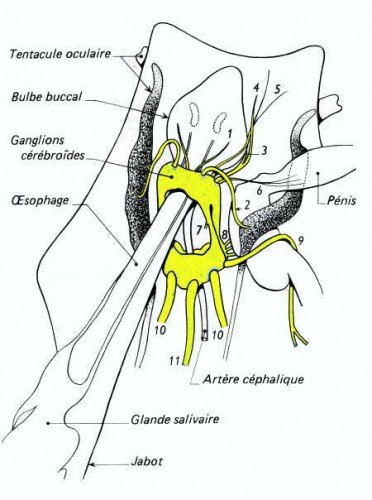
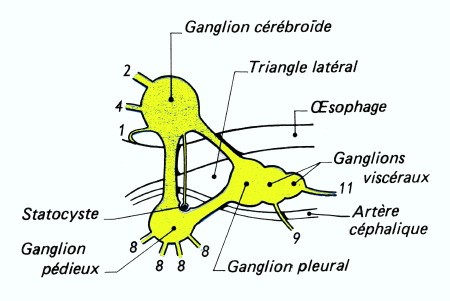
Quant au collier périœsophagien, il montre de chaque côté le connectif cérébro-pédieux accolé au connectif cérébro-palléal ; le triangle latéral a disparu par suite du rapprochement des ganglions palléaux et pédieux. La chaîne nerveuse est très réduite par suite de la remontée des ganglions viscéraux près des ganglions palléaux : elle ne montre pas la torsion en X, caractéristique des Gastéropodes Prosobranches.
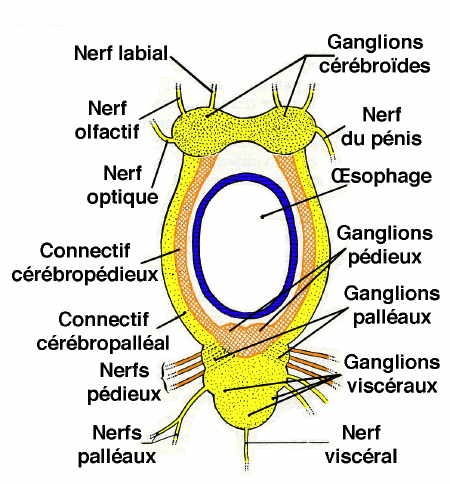
Organes des sens
En dehors des cellules neuro-épithéliales, on trouve :
- deux tentacules antérieurs tactiles et gustatifs ;
- deux tentacules postérieurs présentant, du côté interne, un organe olfactif et, à l'extérieur un œil. Celui-ci est formé par une cupule de cellules pigmentées abritant des cellules sensorielles. Ces organes sont plus sensibles aux rayons infrarouges qu'aux radiations visibles ;
- deux otocystes, logés chacun dans une cavité close près du ganglion pédieux, mais innervés par les ganglions cérébroïdes.

Appareil digestif
La bouche donne accès dans un bulbe buccal ou pharynx. Dans celui-ci, on trouve, en avant et ventralement, une langue formée d'un bourrelet musculaire, rejetée vers l'arrière; au-dessus, une mâchoire chitineuse permet de sectionner les feuilles. Plus en arrière, la radula est un ruban chitineux porté par une saillie ventrale ; sur ce ruban sont fixées des dents chitineuses, orientées vers l'arrière et disposées en rangées parallèles ; leur forme est caractéristique des différentes espèces de Gastéropodes, pour lesquels on peut établir des formules dentaires.


Radula d'escargot (Pomatias elegans) Lumière polarisée
© Christian Gauthier
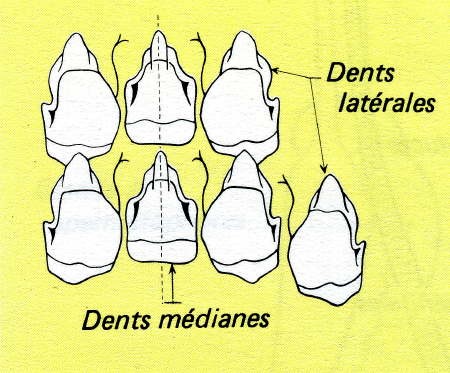
Cette radula prend naissance dans la gaine de la radula, repli de la muqueuse buccale dont les cellules épithéliales ou odontoblastes sécrètent la chitine. La masse qui porte le ruban chitineux est renforcée par un cartilage sur lequel s'attachent des muscles protracteurs de la radula, contenus dans la langue, et des muscles rétracteurs orientés vers l'arrière. Animée de mouvements longitudinaux, la radula fonctionne comme une râpe déchiquetant très finement les végétaux. Cette mastication est facilitée par une salive abondante, la "bave" de l'Escargot ; celle-ci est fournie par deux glandes salivaires importantes, allongées de part et d'autre de l'œsophage et du jabot, et communiquant chacune avec le bulbe buccal par un canal assez long.
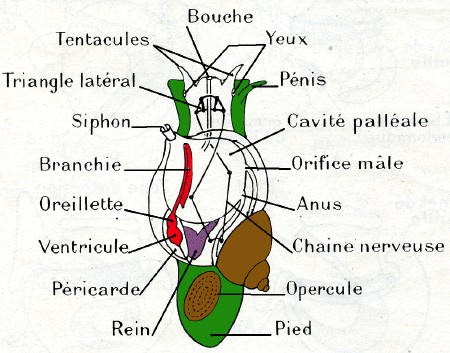
Du bulbe buccal, le tube digestif se continue vers l'arrière par un œsophage qui se renfle ensuite en jabot ; à la base du tortillon, il donne accès à une poche sphérique, l'estomac, entouré par l'hépato-pancréas dont il reçoit les canaux excréteurs. L'intestin, long, comme chez les herbivores, revient vers l'avant pour former un rectum qui débouche par l'anus près du pneumostome, sur le côté droit de l'animal.
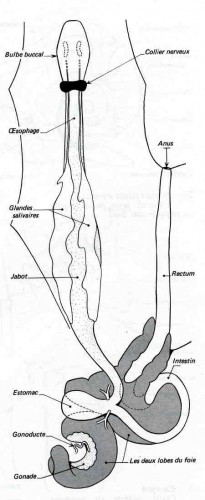
L'hépato-pancréas ou glande digestive sécrète des diastases, en particulier une cytase capable de digérer la cellulose. Il fonctionne également comme un organe de réserve glycogénique. Certaines cellules contiennent du phosphate de calcium : c'est une réserve de calcaire utilisée pour la croissance de la coquille et la formation de l'épiphragme. La glande accumule également du fer et a aussi une fonction excrétrice.
Appareil respiratoire
Il est constitué par la cavité palléale remplie d'air et transformée en poumon ; elle ne communique avec l'extérieur que par le pneumostome. Les vaisseaux sanguins où se fait l'hématose sont contenus dans la paroi du manteau qui forme le toit du poumon. Les échanges gazeux ne se font bien que si l'air de la cavité palléale est humide ; par temps sec, l'animal se rétracte dans sa coquille, ce qui diminue l'évaporation de la pellicule d'eau pulmonaire.
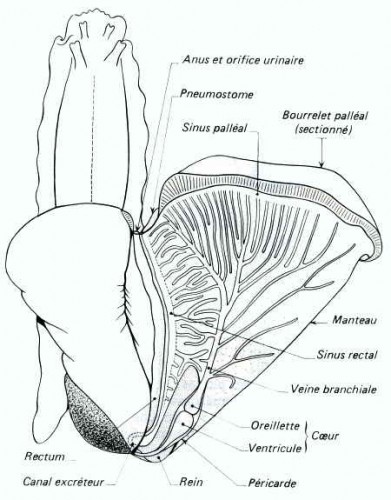
Appareil circulatoire
Le cœur est situé juste en arrière de la cavité palléale ; la position antérieure du complexe palléal résulte de la torsion affectant le développement des Gastéropodes ; de même la dissymétrie résultant de l'enroulement en hélice de la masse viscérale a fait disparaître une oreillette ; celle qui subsiste est devenue antérieure par rapport au ventricule.
Du ventricule se détache une artère se divisant en deux : l'aorte antérieure dessert la tête et donne, en particulier, une artère salivaire, une branche génitale du côté droit, et une branche palléale ; l'aorte postérieure se ramifie surtout dans l'hépato-pancréas.
Des artères, le sang, légèrement bleuté par l'hémocyanine, passe dans un système de lacunes, puis de sinus veineux, surtout développés dans le pied ; c'est l'accumulation du sang dans les sinus pédieux qui provoque la turgescence du pied.
Les sinus veineux convergent vers le rein, puis le sang passe dans le toit de la cavité palléale et, après l'hématose, est ramené à l'oreillette.
Appareil excréteur
L'enroulement de la masse viscérale a fait disparaître un rein ; il n'en subsiste qu'un, appliqué contre le péricarde. Il débute dans cette cavité péricardique et le canal présente une paroi très plissée et glandulaire, richement irriguée par des sinus veineux. Le canal excréteur se dirige vers l'avant et va longer le rectum. L'orifice excréteur est situé près de l'anus, entre celui-ci et le pneumostome.
Appareil génital
Particulièrement complexe, il est hermaphrodite, ce qui constitue un caractère d'évolution parmi les Gastéropodes.

Le spermiducte aboutit à l'orifice génital par une partie terminale renflée à parois musculeuses, le pénis, organe d'accouplement ; le pénis se prolonge postérieurement par une glande étroite et filiforme, la glande du fouet ; sur le pénis, s'attachent les muscles rétracteurs du pénis.
L'oviducte débouche dans un vagin en forme de sac qui s'ouvre également à l'orifice génital ; au fond du vagin, la poche du dard sécrète le dard, pièce calcaire à rôle excitateur ; le vagin est flanqué d'une paire de glandes multifides, très ramifiées, sécrétant un mucus. Enfin, du vagin part un fin canal aboutissant au réceptacle séminal.
Voir la suite l'Escargot 5.
Sources :
- Cours de zoologie de André Guyard
- Boué H., Chanton R. (1958) - Zoologie I. Invertébrés 2. 542 p. Doin Ed. Paris
Documents et photographies non renseignés : André Guyard
17 commentaires
I am interested in knowing where you obtained the image you have used for gastropod torsion (larvae).
It is better than most available
Winston Ponder
Ces croquis sont inspirés de Boué et Chanton, Zoologie I, Invertébrés ** (1958).
peut-on savoir pourquoi l'orifice respiratoire s'ouvre et se ferme au cours de la respiration ?
Le rythme de fermeture et d'ouverture du pneumostome (orifice respiratoire) varie en fonction de l'état d'hydratation de la surface pulmonaire. C'est donc l'état de déshydratation qui initie et module le rythme d'ouverture-fermeture du pneumostome.
Des expériences menées chez la limace Limax maximus, montrent que lorsqu'un individu a perdu 15-20% de son poids corporel initial par évaporation, la fréquence de fermeture du pneumostome, qui est inférieure à 0,5 fermeture/min chez les limaces entièrement hydratées, commence à augmenter.
Cette fréquence s'accroît avec une déshydratation plus poussée, mais la durée moyenne de chaque obturation reste constante. Ainsi, la proportion de temps pendant lequel le pneumostome est fermé augmente. Simultanément, la surface de l'ouverture du pneumostome diminue.
Ce comportement semble être contrôlé en partie par l'osmolalité de l'hémolymphe de la limace et par un peptide étroitement lié à l'arginine-vasotocine (AVT) et l'arginine-vasopressine (AVP).
L'injection de mannitol chez des limaces qui augmente l'osmolalité de l'hémolymphe, peut initier le rythme ouverture-fermeture du pneumostome.
Mais cette injection de mannitol, ne provoque pas la diminution du diamètre de l'ouverture du pneumostome qui est induite par la déshydratation naturelle ou par l'injection d'AVT ou d'AVP.
Cela suggère qu'au moins deux systèmes peuvent être impliqués dans le contrôle global du rythme ouverture-fermeture du pneumostome.
bien tout ce que vous dite mais je croit que vous n'avez pas parler du phénomène de détorsion que subisent également les gastéropodes car dans les lignées de gastéropodes on constate souvent une soete de retour par détorsion et les études menées montrent aujoudhui que ce phénomène survient juste aprés la torsionqui précède l'enroulement dela masse viscérale et que ce phénoméne n'affecte que les lignées les plus évolutives comme les euthyneures (opistobranches et les pulmonés( stylomatophoreset bassomatophores) alors que la torsion n'affecte que les plus primitifs comme les stretoneures. en attendant de vous d'emple explication sur le phénoméne de détorsion ? qu'en pensez vous de ceci? merci
Bonjour,
Relisez plus attentivement le texte qui répond à vos interrogations.
disposz vous des information sur les molusques en général? si faites moi un peu une comparaison de l'appareil respiratoire entre ces différentes classes Ya t-il une coreelation évolutive?
avez vous une institut de recheche? si oui comment fait -on pour s'abonner
Sil vous plait est ce que c'est possible de m'envoyé
comment faire la dissection d'un escargot.
Merci d'avance
DISSECTION DE L'ESCARGOT
Pour la dissection, on choisit Helix pomatia ou Escargot de Bourgogne, qui est l'objet d'un important commerce, et que l'on peut se procurer en toutes saisons en France. Dans le Maghreb, on pourra disséquer le Gros Gris (Helix aspersa maxima).
Morphologie — L'animal est enveloppé d'une coquille à l'intérieur de laquelle il peut se rétracter et s'enfermer, en sécrétant un mucus qui se dessèche à l'air. Ce diaphragme n'a aucun rapport avec l'opercule, situé à la face dorsale et postérieure du pied, qui chez un grand nombre de gastropodes sert à l'obturation de l'ouverture de la coquille quand l'animal se rétracte.
La coquille est contournée en spirale de droite à gauche (enroulement dextre) et forme quatre tours et demi de spires. À sa surface, on remarque des lignes d'accroissement.
L'ouverture de la coquille porte le nom de péristome, l'axe d'enroulement est creux et constitue la columelle, dont la partie supérieure correspond au sommet de la coquille, et la base présente un orifice ou ombilic. L'animal n'adhère à la coquille que par un muscle columellaire.
À l'état d'extension, l'Escargot repose sur le sol, par une large et épaisse masse musculaire : sole plantaire ; en avant est située la tête, présentant une bouche antérieure limitée par une lèvre supérieure, une lèvre inférieure et deux lèvres latérales.
Au-dessus de la tête, se trouvent quatre tentacules : deux petits antérieurs et deux postérieurs plus grands portant les yeux.
Un peu en arrière, et sur le côté droit, est située l'ouverture génitale ; du même côté et contre le rebord de la coquille se trouve un large orifice ou pneumostome faisant communiquer le poumon avec l'extérieur ; enfin sur la marge interne du pneumostome s'ouvrent l'orifice anal et le pore excréteur.
L'animal étant tué en extension, par asphyxie dans l'eau qu'on a fait bouillir et refroidir pour la priver d'air, on enlèvera la coquille en la découpant le long des spires avec de forts ciseaux, puis en détachant l'axe de la columelle.
L'animal dépourvu de sa coquille montre le même enroulement que celle-ci. On remarque, en dessus et en avant, la chambre pulmonaire limitée par le repli du bord du manteau ; sur le côté gauche le cœur transparent, et un peu au-dessus l'organe excréteur de teinte gris-jaunâtre ; tout le reste de la masse du tortillon est constitué par l'hépato-pancréas ou glande digestive, à la surface duquel on distingue les circonvolutions de l''intestin ; la pointe du tortillon est occupée par la glande génitale.
Anatomie. — Les différents temps de la dissection doivent être suivis très exactement afin de pouvoir étaler convenablement les organes :
1° On incisera le rebord du manteau, en introduisant des ciseaux dans l'ouverture du pneumostome ; puis on poursuivra l'incision en longeant par l'intérieur le bourrelet qui borde en avant la cavité palléale, et on continuera l'incision, sur le côté gauche, en passant en dessous et en arrière du cœur et du rein, que l'on voit par transparence. Cette incision superficielle — la membrane palléale et les téguments étant très minces — permet d'ouvrir la cavité pulmonaire et de faire basculer le manteau, le rein et le cœur sur le côté droit de l'animal. L'Escargot sera alors fixé, la sole pédieuse contre le liège de la cuvette.
2° On incise ensuite le plancher de la cavité palléale — pour mettre à découvert la masse viscérale antérieure — en sectionnant, suivant la ligne médiane, le rebord du manteau et toute la région antérieure jusqu'au niveau de la bouche.
3° On rejette à droite tous les organes génitaux, et à gauche le tube digestif.
Pour repérer les organes, se référer aux schémas qui illustrent l'article.
Appareil digestif. — À la bouche fait suite un bulbe buccal à l'intérieur duquel se trouve l'appareil masticateur ou radula. Celle-ci est constituée par une lame portant des dents cornées et s'insérant dans une papille ventrale saillante ; au-dessus de la radula est située une dent cornée
Le bulbe se continue par un œsophage qui se renfle en un estomac fusiforme volumineux.
À la suite de l'estomac vient l'intestin contourné en U et circonvolutionné autour du foie, il se continue par un rectum qui aboutit à l'anus, situé sur le bord du pneumostome.
Les glandes annexes du tube digestif comprennent :
Deux glandes salivaires déchiquetées, blanchâtres, en partie anastomosées entre elles, qui enveloppent l'estomac et la bouche par de fins canaux dans le bulbe ; et un hépato-pancréas volumineux occupant tout le tortillon et s'ouvrant dans le tube digestif entre l'estomac et l'intestin.
Appareil circulatoire et respiratoire. — Le cœur est enveloppé d'un péricarde ; il est formé d'un ventricule postérieur d'où part en arrière l'aorte, et d'une oreillette antérieure où aboutit la veine pulmonaire. Celle-ci se ramifie dans la cavité palléale, qui communique avec l'extérieur par le pneumostome. La cavité palléale joue le rôle de poumon.
Appareil excréteur. — Il n'existe qu'un seul rein, ou organe de Bojanus, situé à côté du cœur et communiquant avec la cavité péricardique. Le rein débouche à l'extérieur, à côté de l'anus par un canal longeant le rectum.
Appareil génital. — L'Escargot est hermaphodite. La glande génitale hermaphrodite est située dans la partie terminale du tortillon ; son conduit hermaphrodite, fin et très flexueux, se dédouble, après avoir reçu les sécrétions d'une volumineuse glande de l'albumen, en un oviducte et un canal déférent ou spermiducte.
Ces deux conduits, l'un flexueux : l'oviducte, l'autre non ondulé : le canal déférent, sont accolés puis après un certain trajet s'isolent l'un de l'autre.
Le canal déférent aboutit à un pénis renflé auquel est fixé un muscle rétracteur et un long flagellum.
L'oviducte vient déboucher dans une poche résistante de couleur nacrée : poche du dard, à laquelle sont annexées deux groupes de glandes : les glandes multifides. La poche du dard contient à son intérieur un spicule calcaire acéré. Au niveau de la poche du dard, se détache de l'oviducte un long
canal récurrent : canal de la vésicule séminale, qui aboutit à un réservoir séminal situé au voisinage de la glande de l'albumen.
Les deux conduits génitaux : pénis et oviducte se réunissent en un vestibule commun qui aboutit à l'orifice génital hermaphrodite. L'accouplement est réciproque, les spermatozoïdes de l'un des individus s'accumulent dans la vésicule séminale de l'autre, où ils séjournent jusqu'au moment de la maturation des ovules.
Système nerveux. — II comprend deux gros ganglions cérébroïdes, unis par une large commissure sus œsophagienne, deux ganglions pédieux situés au-dessous du bulbe et réunis entre eux, et trois ganglions viscéraux en arrière du bulbe dont deux latéraux et un médian accolés les uns aux autres.
Ces divers ganglions sont réunis entre eux par des connectifs cérébro-pédieux, cérébro-viscéraux, et viscéro-pédieux.
Ces divers ganglions et leurs connectifs sont englobés par du tissu conjonctif dont la masse forme un large collier périœsophagien. Pour distinguer ces ganglions il faut enlever l'ensemble du collier et le traiter par l'acide acétique (vinaigre).
Musculature. — En dehors des muscles du pied dont les fibres sont disposées en tous sens et mêlées à de nombreuses glandes à mucus, il existe un muscle columellaire, étalé dans le pied, et unissant l'animal à sa coquille.
bsr je veux savoir svp quel est la glande qui secrète la bave ? et les molécules qui régulent cette sécrétion. Merci
Bonjour,
Les glandes qui sécrètent "la bave" sont les glandes à mucus qui tapissent l'épiderme de l'escargot.
En ce qui concerne la régulation de la sécrétion, j'en ignore les mécanismes.
Mais on peut en avoir une idée en consultant la thèse d'université de Mélanie Gay (2004) concernant la moule Crassostrea gigas dont voici un extrait :
Mécanismes de défense chez la moule Crassostrea gigas (extrait de la thèse d'université de Mélanie Gay, soutenue le 10 décembre 2004, Université de la Rochelle)
La coquille et le manteau constituent une protection mécanique contre les agressions extérieures. En cas de danger détecté par les tentacules disposés le long du manteau, la coquille se ferme. Quand la coquille est ouverte, le manteau fait office de filtre, rejetant les particules trop grosses.
Le système immunitaire des huîtres est non adaptatif puisqu'il est dépourvu des acteurs de la mémoire immunitaire (cellules lymphoïdes et anticorps). Il possède néanmoins des mécanismes de reconnaissance soi/non-soi ainsi que les deux grands types de réponses immunitaires, cellulaire et humorale.
1. Les effecteurs du système immunitaire : les hémocytes
Les hémocytes sont présents dans le réseau sanguin (cœur, artères, sinus veineux, veines) et dans les lacunes sanguines des organes, mais sont également infiltrés dans les tissus. Cependant le lieu et le processus de l'hématopoièse sont encore inconnus (Barreau-Roumiguiere et al., 2003). Malgré quelques légères différences, les auteurs s'accordent à décrire deux grands types d'hémocytes chez les bivalves (Hme, 1999), les granulocytes et les agranulocytes ou hyalinocytes.
Les grandes fonctions des hémocytes sont (Cheng, 1981 ; Feng, 1988 ; Fisher, 1988 ; Hine, 1999) :
o l'agrégation : formation d'amas cellulaires (clou hémostatique) réversibles suite à des stimulations ;
o l'adhésion et le déplacement sur un substrat inerte ou non : phénomènes qui participent avec l'agrégation à la réparation de lésions tissulaires ou de la coquille ;
o la plasticité : par le déploiement de pseudopodes, les hémocytes passent d'une forme sphérique à une forme amoeboïde facilitant l'infiltration et le déplacement ;
o le chimiotactisme : les hémocytes sont attirés par des substances produites par les organismes étrangers ou par l'huître en cas de blessure ou de réaction inflammatoire ;
o la réparation de blessures : par infiltration du site lésé et colmatage de la lésion, différenciation cellulaire (production de fibres de collagène liant les hémocytes agrégés entre eux), phagocytose des débris cellulaires et réparation des épithéliums ;
o la réparation de la coquille par mobilisation et transport de calcium et de protéines vers la lésion, puis dépôt de ces éléments sur le site de la lésion ;
o la sécrétion/excrétion : production de la majorité des enzymes hydrolytiques, molécules cytotoxiques, substances intervenant dans les activités hémocytaires, substances associées à 1'homéostasie, substances de modulation de la réponse immunitaire, du stress ou de l'inflammation ;
o les mécanismes de défense ;
o la digestion et le transport des nutriments.
2. Immunité cellulaire
Elle comprend les mécanismes de défense impliquant les hémocytes eux-mêmes. Les trois phénomènes principaux sont l'infiltration, la phagocytose et l'encapsulation.
L'infiltration conduit à une forte concentration d'hémocytes sur le site de la lésion ou de l'infection. L'issue peut être soit la destruction ou l'isolement de l'organisme étranger, soit la réparation complète du tissu, soit, dans quelques cas extrêmes, la destruction du tissu de l'hôte.
Les principales étapes sont : infiltration du lieu lésé ou infecté par de nombreux hémocytes, formation tfun amas par des hémocytes agrégés, remplacement des tissus endommagés par des hémocytes allongés, dépôt de collagène, élimination des tissus nécrotiques par des hémocytes phagocytaires et restauration de l'architecture normale du tissu (Feng, 1988). À ce jour, l'origine des hémocytes infiltrants n'est pas encore clairement établie, il s'agit soit du recrutement de cellules présentes dans d'autres compartiments, soit d'une multiplication in situ. Les hémocytes infiltrants agissent dans la lutte contre l'agent infectieux et/ou dans la réparation des tissus lésés (Ford et al,, 1993).
La phagocytose est le mécanisme le plus important et le mieux décrit car il est facilement reproductible in vitro (Xue et al., 2001 ; Lambert et al., 2003 ; Goediten & De Guise, 2004). Les différents événements sont similaires à ceux décrits chez les mammifères. La première étape est la reconnaissance de l'organisme comme étranger par l'intermédiaire de récepteurs et de lectines (Olafsen et al., 1992). Après adhésion de la particule étrangère sur l'hémocyte, la membrane cytoplasmique de celui-ci englobe la particule étrangère par contraction du cytosquelette. L'internalisation de la particule dans le cytoplasme de l'hémocyte se fait sous forme d'un phagosome qui fusionne avec un ou plusieurs lysosomes pour former le phagolysosome, la particule est détruite par les enzymes lysosomiales ou par la flambée oxydative. Cette dernière consiste en la production de radicaux libres, formes intermédiaires réactives de l'oxygène, et de peroxyde (l'hydrogène, Les produits de dégradation de la particule sont soit utilisés par la cellule (accumulés dans des vacuoles sous forme de pigment tel la mélanine), soit libérés dans le milieu extracellulaire par l'intermédiaire de vésicules (Feng,1988; Xue,1998).
Différents types cellulaires sont capables de phagocytose. Parmi les hémocytes, les granulocytes présentent la plus forte activité phagocytaire (Hine, 1999 ; Goedken & De Guise, 2004), Les cellules céreuses ou cellules brunes participent également au phénomène de phagocytose sur les sites de lésions et d'infections (Hine, 1999). L'encapsulation concerne les agents pathogènes métazoaires (trématodes, cestodes) de grande taille ne pouvant pas être phagocytés. L'agent infectieux est entouré d'une couche interne constituée de cellules comparables à des fibroblastes et d'une couche externe constituée de matériel fibreux contenant des glycoprotéines et/ou des mucoprotéines et infiltrée par des hémocytes. La destruction du pathogène serait due à la p-glucoronidase (Feng, 1988).
3. Immunité humorale
Il s'agit de l'ensemble des substances luttant contre des agents pathogènes, des toxines ou des polluants (Chu, 1988 ; Xue, 1998 ; Roch, 1999) :
o les enzymes hydrolytiques responsables de la lyse intra ou extracellulaire (lysozyme, p-glucuronidase, phosphatases acide et alcaline, serine protéase, amylase.lipases) ;
o les lectines, protéines pouvant se fixer spécifiquement à des sucres, fonctionnent comme des opsonines en reconnaissant des ligands étrangers et peuvent être induites par la présence de pathogènes (Olafsen et al., 1992) ;
o le système prophénoloxydase-phénoloxydase : la phénoloxydase activée catalyse la réaction de conversion de substances phénoliques en mélanine ; les composés intermédiaires de cette réaction sont bactéricides et interviennent dans les phénomènes de reconnaissance du non-soi (Luña-Gonzalez et al., 2003) ;
o les protéines de stress (heat shock proteins) permettent d'accroître la capacité des cellules à supporter les variations de l'environnement (Boutet et aL, 2003) ;
o la NO synthase catalyse la production d'oxyde nitreux qui intervient en tant qu'agent anti-bactérien et anti-tumoral (Arumugan et al., 2000) ;
o les peroxydases interviennent dans les mécanismes de la flambée oxydative ;
o des molécules cytotoxiques et certaines substances physiologiques telles la transferrine et la lactoferrine inhibent les agents pathogènes (Roch et al., 1996 ; Montagnani et al., 2001 ; Romesland et al,, 2002 ; Gueguen et al., 2003).
À ce jour, aucun peptide anti-microbien n'a été caractérisé chez C. gigas alors que ce type de molécule est décrit dans tout le règne vivant, y compris chez certains mollusques (Mitta et al., 1999 ; Gueguen et al., 2003) et que des activités antibactériennes et antivirales ont été observées dans l'hémolymphe d'huîtres (Hubert et al., 1996 ; Olicard, 2003). Des mécanismes de détoxification mettant en jeu la métallothionine et les glycoprotéines-P transmembranaires ont été mis en évidence chez l'huître pour l'élimination de polluants ou de toxines (Anderson et al., 1999 ; Bard, 2000 ; Boutet et al., 2004).
Bonjour monsieur, je suis actuellement en classe de BCPST nous réalisons un TIPE sur les escargots plus précisement sur la régénérescence des coquilles d'escargot. Nous savons que l'escargot sécrète la sutrutre par le manteau par le biai de glande. Nous aimerions avoir des informations sur ces glandes si vous en avez. merci d'avance pour votre réponse.
Bonjour,
Je n'ai jamais travaillé sur la problématique de la régénération de la coquille de escargots, mais sur celle de la différenciation des gamètes dans cette glande hermaphrodite.
Les escargots sont capables de réparer leur coquille lorsque celle-ci est abîmée. En effet, le mucus d’escargot sécrété par des cellules glandulaires du manteau contient des substances permettant en particulier l'élaboration et évidemment la régénération de la coquille abîmée. Les substances en jeu sont l’allantoïne, le collagène, l’élastine, l’acide glycolique, des vitamines et des antibiotiques.
L’escargot possède la particularité de régénérer ses propres tissus et même sa coquille, par l’intermédiaire de la production de calcaire qu’il obtient de sa nourriture et grâce à l’effet de l’allantoïne.
Les vitamines sont obtenues par l’escargot à travers l’alimentation à base de végétaux riches en vitamines.
L’acide glycolique contribue à l’action régénératrice des tissus endommagés.
Ces propriétés sont exploitées par les laboratoires pharmaceutiques et cosmétiques pour proposer des médicaments de régénération de la peau humaine.
Approfondissez vos recherches bibliographiques sur internet. Bon courage !
Merci beaucoup pour cette réponse, elle va beaucoup nous aider.
Bonne journée.
Bonjour Monsieur,
Serait'il possible de réutiliser votre dessin de l'appareil génital de l'escargot. L'utilisation en serait : Fiche numérique mise en Intranet dans un projet d'outils scolaires à destination des élèves de Suisse romande.
Merci de votre aide, bien cordialement
Nathalie (merci de me répondre par mail)
Bonjour,
C'est d'accord
Les commentaires sont fermés.