
Coraux des Antilles (6)
19/07/2010
Nutrition, calcification, reproduction et développement des coraux de la Province caraïbe
 Coraux et milieu récifal de la Province Caraïbe
Coraux et milieu récifal de la Province Caraïbe
Chapitre VI : Biologie des coraux
par André Guyard
(suite du chapitre V : coraux du récif)
1. Nutrition des coraux
1.1. Sources de nourriture
Les coraux sont capables de capturer des proies planctoniques et d'absorber des substances nutritives dissoutes dans l'eau de mer à travers l'ectoderme. En outre, ils hébergent des algues unicellulaires, les Zooxanthelles qui leur assurent une nutrition photosynthétique.
a. Proies en suspension
À l'aide de leurs tentacules, les polypes capturent les éléments du nanoplancton (microcrustacés et bactéries). Si l'on envisage la totalité de la colonie, on conçoit que l'ensemble des polypes représente une masse considérable de capture. Les proies, paralysées par le venin des nématoblastes, sont dirigées vers la bouche par un film de mucus convergeant grâce à des mouvements ciliaires. D'après Goreau (l956), ce mouvement se fait en moins de 30 s chez Siderastrea siderea. C'est en général la nuit que la capture est active, les polypes étant alors en extension, probablement pour profiter de la migration ascendante nocturne du plancton.
Le cliché ci-dessous réalisé par Douglas Brumley (Institut de technologie du Massachusetts) a été classé à la 6ème place au concours des plus belles photos microscopiques du Nikon Small World 2014. Il matérialise les courants d'eau engendrés par le mouvement des cils.
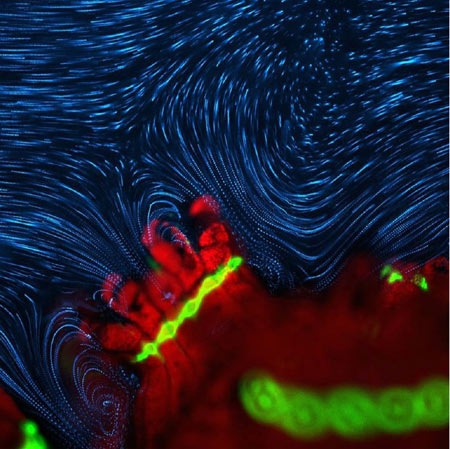
Écoulement d'un fluide autour d'un polype corallien
© Dr. Douglas Brumley / Courtesy of Nikon Small World
b. Absorption de matériel en solution
Les matières organiques en solution (acides aminés, glucides et macromolécules) sont directement absorbées par l'ectoderme dont les cellules présentent des microvillosités favorables à la pinocytose. En outre, les ions phosphate, hydrogénocarbonate, calcium, assimilables par le tégument, permettent la squelettogenèse calcaire.
c. Symbiose avec les Zooxanthelles
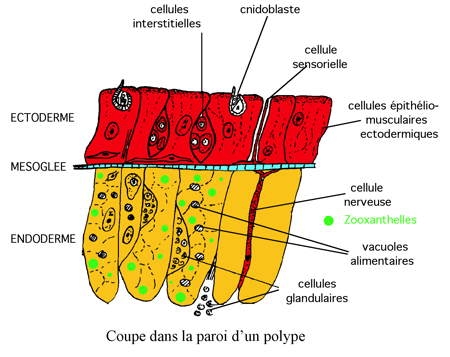
Les Zooxanthelles, algues vertes unicellulaires (Dinoflagellés) situées dans les cellules endodermiques des coraux hermatypiques vivent en symbiose avec le polype. Leur importance n'est pas négligeable puisque le rapport protéines animales/protéines algales est d'environ de 1. Ce sont des algues photosynthétiques qui vivent et se divisent à l'intérieur des cellules de l'hôte. C'est sur cette symbiose que repose la productivité de l'écosystème récifal (voir plus loin chapitre IX).
La source d'énergie nécessaire à la photosynthèse étant la lumière, les coraux hermatypiques ne peuvent prospérer à une profondeur supérieure à 70 mètres en Mer Caraïbe. D'autre part, ils ne tolèrent pas une température inférieure de 21 °C ni l'abaissement de la salinité, ni une turbidité élevée.
À faible profondeur où la lumière est abondante mais où l'énergie des vagues est forte, les coraux développent des squelettes aux branches robustes. À une profondeur plus grande ou dans les zones où l'éclairement est moindre, les colonies se présentent sous la forme de plateaux horizontaux et chaque polype abrite des Zooxanthelles plus nombreuses.
Les Zooxanthelles sont contenues dans des vésicules closes des cellules gastriques du polype-hôte. Le corail exerce un contrôle sur la population des Zooxanthelles et expulse par exocytose les algues les plus âgées, moins actives du point de vue métabolique.
L'association Corail-Zooxanthelles est un bel exemple de symbiose. chaque organisme trouve son compte dans cette coexistence.
Goreau et Goreau (1971) marquant au 14C radioactif les nutriments ingérés par les polypes, ont montré que, le jour, les Zooxanthelles absorbent ce 14C et l'assimilent par photosynthèse en matière organique. Une partie de cette matière organique est libérée par les algues au profit de l'hôte sous forme de glucose, de glycérol et d'acides aminés. Le Corail profite également de l'oxygène rejeté par la photosynthèse.
Étant donné la simplicité d'organisation des coraux, l'excrétion des déchets minéraux solubles n'est pas confiée à un appareil particulier et se fait pas simple diffusion dans la cavité digestive. Ces ions PO43-, NO3-, SO42- et NH4- sont récupérés directement par les Zooxanthelles qui profitent de ces nutriments particulièrement rares en eau tropicale.
Ainsi, la demande photosynthétique des Zooxanthelles aboutit à un recyclage des déchets des coraux et leur transformation en matière organique. De sorte que les coraux sont capables :
- de se nourrir activement aux dépens du plancton ;
- de prélever les sels minéraux nutritifs dans l'eau de mer ;
- d'absorber les substances chimiques libérées par les Zooxanthelles.
Ils participent donc à différents échelons de la chaîne alimentaire. Ils sont producteurs primaires grâce à leurs Zooxanthelles, consommateurs primaires et secondaires. La complexité de ce réseau trophique réduit leur dépendance vis-à-vis des sources de nourriture.
1.2. Digestion et assimilation
La taille des proies dépasse souvent largement la taille des cellules digestives de sorte qu'une digestion extra-cellulaire précède la phagocytose et la digestion intra-cellulaire au niveau de l'endoderme. La présence de proies dans la cavité gastro-vasculaire stimule la sécrétion d'enzymes protéolytiques. L'excrétion se fait également dans la cavité digestive et la bouche sert alors d'anus. Il faut souligner l'importance des mésentéries, ces cloisons radiaires qui accroissent d'une façon considérable la surface digestive. Certaines espèces ont la faculté de dévaginer leurs filaments mésentériaux et de digérer à l'extérieur de la cavité gastrique des proies de taille supérieure au diamètre de la bouche. Cette propriété est d'une grande importance comme nous le verrons plus loin pour la compétitivité entre espèces. Grâce à l'intercommunication par les canaux du cœnosarque des cavités gastriques des différents polypes, la capture d'une proie par un individu profite à toute la colonie.
2. Calcification des coraux
Le squelette des coraux est constitué d'aragonite, une forme cristalline fibreuse de CaCO3. Des cellules spécialisées, les calicoblastes absorbent l'ion Ca2+ de l'eau de mer et le transfèrent vers les sites de calcification par diffusion et pompage actif. Or, le taux de calcium intra-cellulaire doit être faible pour que les cellules puissent fonctionner normalement. Böhm a montré que chez les coraux, bien que la teneur des tissus en calcium soit du même ordre de grandeur que celle de l'eau de mer, ces animaux ont une concentration faible en Ca2+ libre du fait que la plus grande partie du calcium est liée aux membranes ou à des molécules organiques. Ce calcium lié se renouvelle très rapidement et se trouve précipité en cristaux minuscules de CaCO3 contenus dans des vésicules qui, par exocytose, les déversent à travers la membrane plasmique vers les sites de calcification (N. Goreau). L'ion CO32- provient du CO2 issu de la respiration grâce à l'anhydrase carbonique.
Nous verrons plus loin que la vitesse de croissance des coraux dépend de l'intensité lumineuse ambiante grâce à l'intervention des Zooxanthelles. Ces algues stimulent le taux de calcification de deux façons :
- en fixant le CO2, ce qui accroît le pH du cytoplasme et la concentration des ions CO32-. Par précipitation des CO32- en excès sous forme de CaCO3 insoluble, le polype ramène son pH à un niveau normal et en même temps, contribue à l'édification de son squelette calcaire.
- en augmentant la quantité d'énergie libre (ATP) disponible pour assurer le transport actif des ions Ca2+, les Zooxanthelles stimulent également la calcification. Ainsi les algues travaillent en synergie avec l'anhydrase carbonique pour stimuler la squelettogenèse.
Les coraux fournissent un bel exemple de symbiose au sens strict, c'est-à-dire d'une coopération avec intérêts mutuels entre cellules animales - le polype - et cellules végétales - les Zooxanthelles -.
3. Reproduction et développement des coraux
3.1. La reproduction sexuée
La dissémination des espèces est assurée par la reproduction sexuée. La formation de larves vagiles permet aux espèces de se fixer à une distance plus ou moins grande de la colonie-mère.
Les coraux sont en général hermaphrodites. Les mésentéries abritent les organes mâles et femelles, sortes de capsules primitives qui renferment les cellules sexuelles : spermatozoïdes et ovules. Les gamètes se forment à partir des cellules interstitielles ectodermiques qui ont conservé un aspect embryonnaire indifférencié. Les testicules et ovaires se trouvent sur le même polype ou au contraire chez des polypes distincts de la même colonie. Une possibilité d'autofécondation est envisageable chez des espèces comme Manicina areolata, Diploria labyrinthiformis et Favia fragum chez qui on rencontre des polypes produisant simultanément ovules et spermatozoïdes. D'autres espèces montrent des colonies unisexuées ce qui suggère un hermaphrodisme successif avec changement de sexe. Chez Acropora palmata, Isophyllia sinuosa, Montastrea cavernosa, Siderastrea radians, seul le sexe femelle a été décrit, alors que chez d'autres espèces, seul le sexe mâle a été découvert. Il est probable qu'il existe des colonies de l'autre sexe et que la fécondation croisée soit la règle.
La fécondation a lieu le plus souvent à l'intérieur de la cavité gastro-vasculaire, le sperme étant charrié par l'eau de mer d'une colonie mâle vers une colonie femelle jusqu'aux polypes contenant les ovules matures. Cependant chez Manicina areolata et Favia fragum, l'expulsion des ovules et du sperme permet d'envisager l'hypothèse d'une fécondation externe.
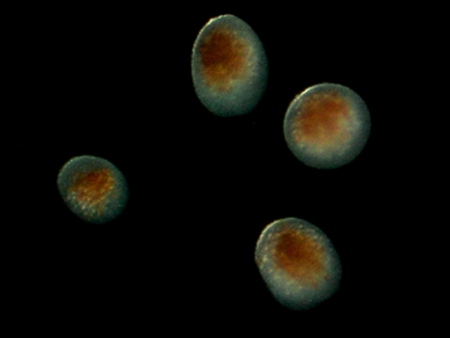
Au bout de quelques jours, par segmentation, l'œuf donne une larve planctonique ciliée, la planula qui mesure entre 0,5 et 2,5 mm. Elle est immédiatement contaminée par les Zooxanthelles qui s'installent au niveau de l'endoderme. À l'issue de la segmentation, la planula est constituée d'un épithélium ectodermique entourant un massif endodermique qui se creusera plus tard d'une cavité gastrique. La larve, expulsée par l'ouverture buccale du polype-mère s'intègre au plancton et mène une vie pélagique en nageant par saccades et mouvements tourbillonnaires grâce à ses cils ectodermiques.
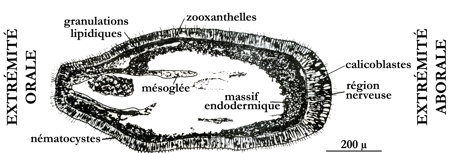
Coupe histologique d'une larve planula
Chez Millepora qui est un Hydrocoralliaire, les cellules germinales se développent dans les tissus de l'animal à partir des cellules interstitielles et migrent vers certains polypes qui se transforment en de petites méduses de 0,5 mm de diamètre et qui portent soit les spermatozoïdes, soit les ovules. Devenues libres, ces méduses lâchent leurs produits sexuels dans la mer où se produit la fécondation.
La reproduction sexuée est un phénomène saisonnier ou continu selon les espèces et les lieux. Agaricia fragilis aux Bermudes et Manicina areolata à Tortugas (Floride) ont un cycle annuel et ne libèrent des larves qu'en été, alors qu'à Hawaii, les larves de Pocillopora sont libérées chaque mois au moment de la nouvelle lune en été et au moment de la pleine lune en hiver. Le nombre d'œufs ou de larves produit par un adulte est variable selon l'espèce. On a pu dénombrer 600 planulas issues d'une colonie de 1850 polypes dans une espèce à petits polypes et une centaine d'ovules par polype dans une espèce à polypes de grande taille.
La vie larvaire est souvent très courte et la fixation a lieu la plupart du temps dans les deux premiers jours.
La distance parcourue pendant cette période dépend évidemment de la vitesse des courants. La fixation est individuelle et chaque larve qui réussira sa fixation au substrat redonnera une colonie. Cependant des larves de même espèce se fixant côte à côte peuvent former une colonie unique.
Une fois fixée sur un substrat solide, la larve subit de profondes modifications. Elle perd ses cils et se transforme peu à peu en un polype solitaire qui commence à sécréter les premiers éléments de son squelette. Peu à peu, le polype grandit, son squelette croît et se diversifie. Les espèces à polype solitaire sont peu nombreuses. Citons Phyllangia americana, Astrangia solitaria, Scolymia lacera et Scolymia cubensis. Chez les espèces coloniales, lorsqu'il a atteint sa maturité, le polype ébauche des bourgeons et donne une colonie par multiplication asexuée. La maturité sexuelle est atteinte en un temps variable : 8 mois chez Favia, 10 ans chez Fungia, polype solitaire indo-pacifique.
La colonisation de substrats solides par les larves coralliennes fait de ces animaux des organismes pionniers dans la conquête de fonds encore inhabités. Cette colonisation s'opère parallèlement à la dissémination de l'espèce par des fragments de colonies détachés accidentellement de la colonie-mère et qui permettent l'expansion de l'espèce par bouturage.
Malgré tout, l'existence de ces deux modes de dissémination n'explique pas la présence de coraux dans des îles éloignées de la Province Caraïbe comme les Bermudes par exemple. Il faut alors envisager l'hypothèse d'une fixation de larves sur des objets flottants et la création de colonies mobiles qui pourraient aller s'échouer sur les îles. Le Gulf Stream, issu de la Mer Caraïbe et qui baigne les Bermudes, serait responsable de la présence des espèces antillaises à cette latitude.
3.2. La reproduction asexuée
Ce mode de reproduction est la règle chez ces êtres polypiformes. Elle se manifeste par le bourgeonnement et la scissiparité.
Le bourgeonnement est le phénomène le plus répandu. Le bourgeon est une protubérance qui se forme sur la paroi des individus marginaux de la colonie et qui comprend les trois couches fondamentales : ectoderme, mésoglée et endoderme entourant un diverticule de la cavité gastrique.
Dans le cas de la scissiparité, une fissure longitudinale apparaît dans les polypes marginaux, les deux moitiés s'écartent et se complètent ultérieurement tout en restant unies par leur base.
L'individualisation plus ou moins poussée des polypes et l'orientation des bourgeons déterminent la forme de la colonie.
- Si les bourgeons sont bien séparés de la mère et naissent à l'extérieur du calice (bourgeonnement extracalicinal) formant un angle aigu par rapport à l'axe longitudinal, la colonie se ramifie et sera arborescente.
- Si le bourgeonnement se produit au bord du calice, la colonie résultante sera plus touffue, voire massive, les polypes étant plus serrés. Si les bourgeons sont très rapprochés, la colonie se développe en nappe.
- Si le bourgeonnement est intracalicinal, les polypes sont coalescents et les ouvertures gastriques dessinent des vallées cernées de collines longues et sinueuses formant des circonvolutions (coraux-cervelles). La colonie aura alors une forme massive, foliacée, encroûtante, sphérique comme un crâne ou en plaque comme une pile d'assiettes.

Conclusion
La coordination parfaite entre polypes d'une même colonie réalise un individu autonome au niveau de la colonie. L'individualité de chaque polype se fond dans l'individualité de la colonie grâce à
- une coopération nutritionnelle réalisée par les échanges digestifs au niveau des canaux de communication creusés dans le cœnosarque ;
- un squelette commun où la fusion atteint son maximum chez les coraux-cervelles ;
- une sexualité commune ; c'est l'ensemble de la colonie qui est mâle, femelle ou hermaphrodite ;
- un système nerveux synchronisé ; l'excitation d'un polype entraîne l'excitation des autres.
En outre, chez les Hydrocoralliaires, les polypes tendent à se spécialiser. On rencontre des individus nourriciers, les gastrozoïdes entourés d'individus prédateurs ou défensifs, les dactylozoïdes ainsi que des individus reproducteurs, les gonozoïdes. Cette coopération entre polypes, cette spécialisation des polypes font de la colonie la véritable unité biologique qu'on peut considérer comme un super-individu.
La suite : croissance et répartition des récifs coralliens antillais.
Sources :
Guyard A. (1979). Coraux des Antilles. Le corail et son environnement. 113 p. 30 diapositives. CDDP Guadeloupe Ed.
Zlatarski V. N., Martinez Estatella N. (1982) Les Scléractiniaires de Cuba. 472 p. Acad. bulg. Sci. Ed.
Intérêt thérapeutique des coraux (ajout du 22 novembre 2017)
En savoir plus sur https://www.lesechos.fr/16/03/2016/LesEchos/22151-116-ECH...
Les commentaires sont fermés.