
Structure des communautés pisciaires lacustres
30/01/2013
Structure des communautés pisciaires lacustres
par Daniel Gerdeaux
I. SPÉCIFICITÉ DES PEUPLEMENTS PISCIAIRES LACUSTRES
Les lacs constituent des habitats spécifiques dans les réseaux hydrographiques. Ils peuvent être considérés comme des «habitats insulaires» car ils constituent toujours des entités aux caractéristiques écologiques très distinctes des milieux courants avec lesquels ils sont, le plus souvent, connectés bien qu'ils aient été normalement colonisés à partir des rivières et fleuves.
Les périodes de colonisation sont en général brèves et suivent immédiatement les événements géologiques qui établissent des relations avec d'autres milieux aquatiques contenant des poissons. Après l'épisode de colonisation au cours duquel s'est constituée la faune piscicole initiale, la richesse spécifique peut être modifiée par des interactions interspécifiques ou des interactions entre l'environnement lacustre et les espèces présentes.
De plus, si le temps est suffisamment long et les biotopes lacustres variés, une colonisation additionnelle, une spéciation à l'intérieur du lac ou dans des milieux connexes peuvent accroître la diversité spécifique dans le lac.
La distribution des poissons d'eau douce lacustre est un des meilleurs exemples du rôle important de l'histoire géologique en biogéographie. Les lacs des zones boréales procèdent d'une formation assez récente lors de la fin des glaciations alors que dans les zones tropicales la plupart des lacs sont de formation beaucoup plus ancienne. Ces éléments sont à prendre en compte pour comprendre la structure spécifique des peuplements piscicoles des lacs.
1. Richesse spécifique suivant les lacs
Une analyse sommaire de la richesse spécifique des peuplements pisciaires lacustres semble montrer que la latitude est un élément important de la richesse spécifique. En effet, les lacs arctiques contiennent au plus 25 espèces et le plus souvent quelques unes seulement, voire une seule, alors que les lacs tempérés en contiennent jusqu'à 130 et les lacs tropicaux plus de 250. L'importance de la latitude est en réalité beaucoup plus faible quand on analyse le phénomène comme l'ont fait Barbour & Brown (1974). La régression multiple opérée entre la diversité spécifique et des variables du milieu pour 70 lacs répartis dans le monde entier, montre que 30,5% de la variabilité sont expliqués par la surface du lac alors que 3,5% le sont seulement par la latitude (fig. 1).
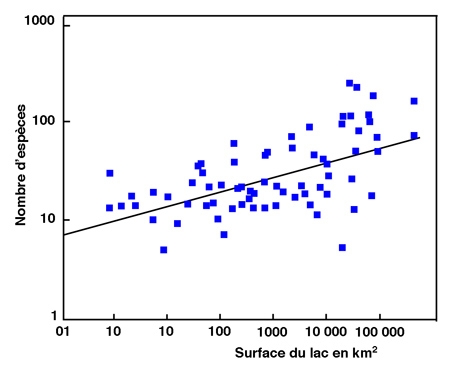
Figure 1 : relation entre l'aire d'un lac et le nombre d'espèces de son peuplement piscicole pour 70 lacs du monde (d'après Barbour & Brown 1974).
La longueur relative des berges et l'altitude du lac viennent ensuite pour expliquer l'essentiel de la variabilité observée. Ceci met bien en évidence les facteurs déterminants de la richesse spécifique.
Un grand lac de basse altitude avec un contour digité offre une grande diversité de biotopes relativement stables qui vont permettre la colonisation de nombreuses espèces ou la spéciation de nouvelles qui vont occuper les milieux variés.
Dans un petit lac de moyenne altitude, la colonisation est plus limitée, les variations du milieu sont plus fortes. Cela ne facilite pas la spéciation. Ce n'est donc pas réellement à cause de leur latitude que les grands lacs tropicaux présentent une grande diversité spécifique mais à cause de leur taille, de leur diversité de biotopes et de leur stabilité. En outre, l'isolement géographique prolongé a facilité une forte spéciation (voir l'endémisme des lacs Baïkal, Tanganyika et Malawi). Dans les lacs arctiques, on trouve à l'opposé des peuplements monospécifiques : le Char Lake (Canada, 75°N, 95°W) ne contient que de l'omble. D'autres lacs, comme les lacs de montagne ne renferment naturellement aucune espèce car la colonisation n'y a pas été possible, les cascades empêchant les immigrations.
L'âge et la taille d'un lac sont donc les deux éléments principaux à prendre en compte pour expliquer sa richesse spécifique.
2. Évolution temporelle de la structure spécifique du peuplement ichtyologique
Un lac évolue au cours du temps ainsi que son peuplement piscicole. L'évolution naturelle se produit à des échelles de temps très longues. L'échelle naturelle la plus petite est celle des lacs formés lors de la dernière glaciation qui datent de moins de 30 000 ans. Par contre, les effets anthropiques provoquent des évolutions rapides à l'échelle de la décennie. Les réservoirs constituent des exemples d'évolution rapide.
Dans les lacs subalpins européens, le peuplement piscicole s'est installé après la dernière glaciation. Des espèces d'eau froide ont pu se maintenir dans ces lacs en profitant de l'existence d'une couche d'eau froide permanente en dessous de la thermocline. Il s'agit de Salmonidés qui furent sans doute les premiers poissons à coloniser les lacs libérés par la fonte des neiges. Les deux principaux genres sont les ombles (Salvelinus) et les corégones (Coregonus).
À l'origine, toutes les espèces étaient probablement migratrices comme le saumon aujourd'hui. Dans les régions boréales, on trouve encore des espèces d'ombles et de corégones qui migrent en mer pour effectuer leur croissance et remontent en rivière pour frayer. Les lacs subalpins ont constitué des refuges et une spéciation a pu alors s'effectuer, en particulier chez les corégones. Ils étaient peuplés de deux à quatre espèces de poissons qui disposaient de différents biotopes à l'intérieur d'un même lac. La thermocline a sans doute constitué une barrière sélective. On trouve ainsi des corégones benthiques souvent petits, avec un régime alimentaire orienté vers la faune du fond et des corégones pélagiques plus gros, planctonophages. Les premiers se reproduisent en hiver ou en été suivant les espèces alors que les seconds ne se reproduisent qu'en hiver. La température de la zone benthique de ces lacs n'atteint jamais un seuil limite pour la reproduction des poissons les peuplant alors que les poissons pélagiques ne trouvent des eaux froides propices à leur reproduction qu'en hiver. Ainsi trouvait-on au début du siècle différentes espèces de corégones dans les lacs subalpins.
La disparition de certains corégones benthiques au début du siècle a été attribuée à une trop forte pression de pêche, mais des travaux plus récents ont montré que certaines disparitions ou diminutions sont à attribuer à la modification de l'état trophique des lacs plus qu'à la surexploitation directe. Quand un lac est eutrophisé, le taux d'oxygène dans les eaux profondes baisse par consommation de l'oxygène lors de la dégradation de la matière organique produite en excès. De plus, une sédimentation accrue conduit au colmatage des fonds sableux ou graveleux constituant les frayères des ombles et des corégones dont les œufs restent en incubation pendant 6 à 8 semaines. La survie des embryons est compromise et le recrutement n'est plus suffisant pour assurer le maintien d'une population soumise, par ailleurs, à une forte pression de pêche. Les individus qui survivent trouvent alors un stock de nourriture d'autant plus grand qu'ils sont peu nombreux à la partager. La croissance individuelle est alors importante. Si la réglementation de la pêche n'est pas changée, la taille minimale de capture ne permet plus une espérance de vie du poisson suffisante pour une bonne reproduction.
On a observé ce phénomène dans le lac Léman entre 1950 et 1980. La teneur en phosphates est passée de 15 mg/l à près de 90 mg/l. La production planctonique disponible comme nourriture a augmenté dans des proportions semblables et les corégones ont grandi plus vite. En 1950, un corégone atteignait 35 cm de long et sa maturité au cours de sa 4ème année de vie au moins alors qu'en 1980 il fait cette taille à la fin de sa deuxième année et est mature l'année suivante. Il s'ensuit rapidement un déséquilibre avec diminution voire disparition de la population de corégones. Parallèlement, les espèces d'eau plus chaude peuvent se développer. Les espèces qui se reproduisent quand la température dépasse 10°C comme la perche ou 19-20°C comme beaucoup de Cyprinidés, sont moins tributaires de la qualité de l'eau pour l'incubation de leurs œufs qui dure moins d'une semaine. Les lacs eutrophes ont donc un peuplement piscicole dominé par ces espèces, qui seront alors défavorisées si le lac redevient mésotrophe quand les mesures de protection de la qualité des eaux sont prises en limitant les apports nutritifs aux lacs.
Dans certains cas, les espèces ajustent leur cycle biologique aux nouvelles conditions du milieu. Dans le lac de Walenstadt, en Suisse, un corégone benthique se reproduisait en été. Actuellement, I'eutrophisation l'oblige à vivre moins profondément dans le lac et, dans ces conditions, il ne trouve plus de conditions thermiques favorables à la reproduction estivale (Ruhlé 1988). Sa période de frai s'est progressivement décalée vers l'automne. La souplesse évolutive des corégones a permis cet ajustement mais cet exemple ne doit pas être généralisé. Le plus souvent, la structure du peuplement est fortement modifiée par l'eutrophisation. Des espèces peuvent disparaître, comme les corégones et l'omble, d'autres prolifèrent.
Dans les réservoirs, cette évolution est encore beaucoup plus rapide et spectaculaire. Le fait de barrer un cours d'eau modifie fortement le milieu. Les espèces d'eau courante disparaissent très rapidement. La mise en eau de surfaces terrestres apporte une grande quantité d'éléments nutritifs qui facilite d'autant plus le développement des espèces bien adaptées aux eaux dormantes que sont la plupart des Cyprinidés.
Le point de rupture du cycle biologique des espèces d'eau courante est presque toujours la reproduction. Les espèces rhéophiles déposent leurs œufs dans les sédiments, sables et graviers alors que les autres espèces pondent sur des substrats comme les herbes ou font des nids qu'elles protègent. La faible exigence de certaines espèces est un atout majeur dans ces conditions. Le gardon est un bon exemple de ces espèces ubiquistes parce que capables de se satisfaire de substrats variés pour la ponte alors que le brochet ne se reproduit que dans des conditions précises. Le gardon peut déposer ses œufs aussi bien sur des cailloux, des herbes, des racines, ou même sur des débris de plastique, alors que le brochet ne développera de comportement reproducteur que s'il rencontre une zone d'herbiers propices.
Un peuplement piscicole est ainsi la résultante des exigences écologiques de chaque espèce qui le compose plutôt que l'intégration de populations constituant une entité fonctionnelle dans l'écosystème. Ceci est valable pour les lacs tempérés, on ne peut pas dire la même chose pour les lacs tropicaux dans lesquels la spéciation a permis une diversification importante et un fonctionnement certainement plus intégré. Les exigences alimentaires très peu marquées chez la plupart des espèces des milieux tempérés permettent facilement des ajustements des peuplements. La truite de lac a un régime alimentaire ichtyophage de préférence. Les jeunes perches constituent la proie préférentielle quand elle est présente, mais à défaut, la truite s'attaque aux jeunes Cyprinidés. En l'absence de poissons, la truite s'alimente sur le compartiment zooplanctonique du lac, et cette nourriture crustacéenne, (riche en caroténoïdes), colore sa chair en rose.
3. Aménagements des peuplements par l'homme
La faible richesse spécifique des peuplements piscicoles lacustres des régions tempérées incite l'homme à des introductions d'espèces pour augmenter la productivité du milieu en poisson. La plupart des modifications ont commencé à la fin du XIXe siècle avec le développement de la pisciculture et la maîtrise de la reproduction artificielle des Salmonidés. Auparavant on ne connaissait pas beaucoup d'exemples de propagation d'espèces exceptée celle de la carpe qui a été réalisée dès l'époque romaine. En effet, on ne pouvait alors que transporter des poissons et non des œufs. Seules des espèces robustes comme la carpe, la tanche par exemple pouvaient faire l'objet de transports importants. Mais dès que l'on a pu transporter des œufs, de grands projets d'introduction d'espèces ont été élaborés. En réalité, il n'y a pas de miracle à attendre d'une introduction mais pas de catastrophe non plus. Quand une chaîne trophique est incomplète, il est logique de penser qu'en la complétant on peut espérer augmenter la production du compartiment ichtyologique sans changer profondément la structure du peuplement présent.
II. STRUCTURE SPATIO-TEMPORELLE DES PEUPLEMENTS PISCIAIRES
La structure thermique d'un lac évolue chaque année suivant le même schéma. Le gradient thermique vertical se met en place progressivement à partir du printemps, se déstabilise à l'automne et disparaît en hiver pour un lac monomictique. Les poissons ont des préférenda thermiques différents suivant les espèces et l'âge qui conduisent les individus à se positionner dans des zones précises au cours d'une année.
1. Occupation de l'espace dans un grand lac au cours d'une année
Pour simplifier, nous considérerons un peuplement lacustre réduit à six espèces : omble chevalier, corégone, perche, gardon, tanche et lotte. Ces espèces ont des exigences écologiques contrastées.
En hiver, la masse d'eau est froide et homogène. Le métabolisme des poissons est très réduit excepté pour les ombles et les corégones poissons d'eau froide qui se reproduisent en décembre : les premiers sur des graviers profonds (40-80 m), les seconds sur des fonds sableux littoraux. Les gardons adultes occupent, en bancs serrés, une zone profonde entre -30 et -50 m, mais se tiennent à proximité du fond. Les jeunes gardons de l'année s'abritent par contre en zone littorale en bancs très importants. Ce comportement est sans doute lié à la prédation. Les perches se tiennent encore plus profondément sur des fonds allant jusqu'à 80 m. Les jeunes perches occupent les mêmes zones et sont l'objet de la prédation par les lottes. En l'absence d'alevins de perches, les lottes remontent dans la zone littorale à la recherche des œufs de corégones ou descendent plus profond sur les frayères à omble (fig. 2). L'occupation de l'espace peut en effet varier en fonction des ressources alimentaires qui ne sont pas toujours les mêmes d'une année à l'autre.
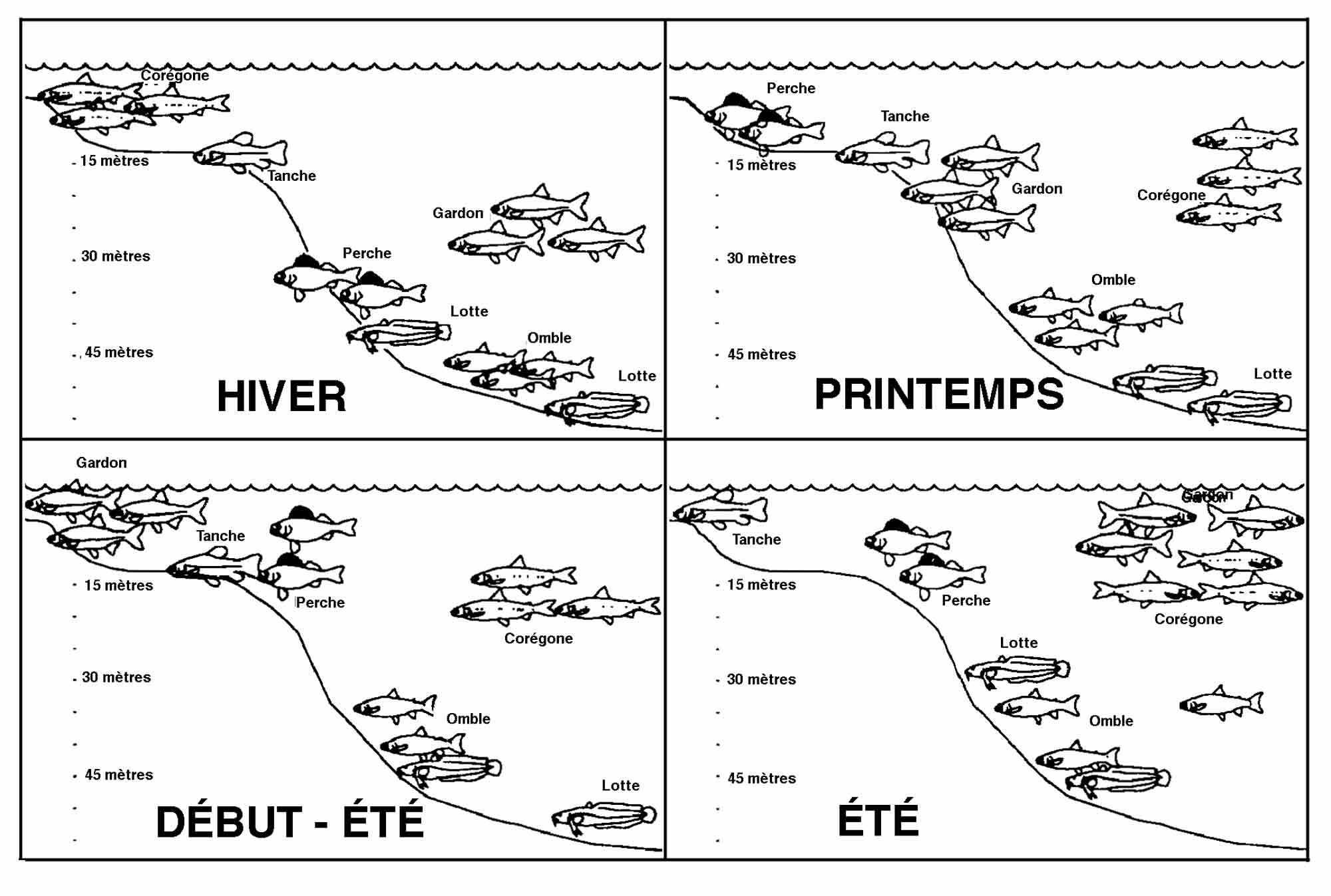
Figure 2 : schémas de la structure spatiale au cours des saisons d'un peuplement piscicole fictif de six espèces adultes en lac profond (d'après Gerdeaux, 1995)
Au printemps, le zooplancton est encore assez rare et les corégones se nourrissent en partie sur le benthos en bordure du mont et les ombles plus profondément. La lotte qui se reproduit en mars, recherche des fonds propices vers 60 à 100 m. La perche se rapproche du littoral pour frayer en mai. Le gardon reste pélagique et s'approche de la couche épilimnique qui se réchauffe légèrement. La tanche reste en zone littorale après son hibernation dans des zones marécageuses à hélophytes.

Figure 3 : échosondage de poissons au lac Léman en septembre, montrant la stratification verticale des poissons. Les petits gardons se tiennent à proximité de la surface, les perches en dessous (vers 15 m), les gros gardons sont à la limite de la thermocline (vers 30 m). Une capture au chalut a permis de vérifier ces identifications. Les échos proches du fond sont probablement ceux d'ombles chevaliers.
En été, la thermocline compartimente verticalement le milieu, mais ne constitue pas une barrière (fig. 3). Les ombles et surtout les corégones la franchissent parfois au cours d'un nycthémère. Les ombles restent toujours à proximité du littoral alors que les corégones se tiennent dans la partie pélagique ainsi que les gardons après leur reproduction, en juin, le long des berges. La perche adulte reste sur le mont au-dessus de la thermocline alors que les alevins après un mois passé au littoral gagnent la pleine eau. La lotte est toujours sur le fond entre 20 et 60 m et la tanche qui a une reproduction fractionnée reste tout l'été dans les zones littorales leur reproduction, en juin, le long des berges. La perche adulte reste sur le mont au-dessus de la thermocline alors que les alevins après un mois passé au littoral gagnent la pleine eau. La lotte est toujours sur le fond entre 20 et 60 m et la tanche qui a une reproduction fractionnée reste tout l'été dans les zones littorales enherbées.
À l'automne, quand la thermocline commence à se déstabiliser, on retrouve une situation proche de celle observée au printemps, mais le zooplancton est suffisamment abondant pour que le corégone et le gardon restent plus au large qu'au printemps.
2. Importance de la répartition spatiale dans les relations inter- et intraspécifiques
Le positionnement des différentes espèces dans la masse d'eau en fonction principalement de la structure thermique diminue les relations entre les différents composants du peuplement piscicole. Les gardons et corégones sont planctonophages en été. Ils partagent ainsi les mêmes ressources trophiques durant une période qui va de mai à octobre (proies principales ingérées : Daphnia sp., Leptodora kindtii et Bytotrephes longimanus. Les recouvrements de leur niche alimentaire durant cette période atteignent des valeurs maximales mais les proies consommées sont alors abondamment représentées dans le milieu. De plus, sans doute en relation avec l'occupation de l'espace, il existe un décalage dans le passage d'un type de proie principale à l'autre au cours de l'été. Les corégones consomment Leptodora kindtii et Bytotrephes longimanus beaucoup plus tôt, beaucoup plus longtemps et en quantité plus importante que les gardons (Ponton & Gerdeaux 1988). Il n'y a pas ainsi de compétition alimentaire importante entre ces deux espèces. Dans la pêche professionnelle orientée sur les corégones, les deux espèces sont capturées simultanément deux mois en moyenne dans l'année.
En outre, pour une même espèce, on constate souvent que l'occupation de l'espace n'est pas la même pour chaque phase du développement. Les jeunes poissons occupent plutôt les couches superficielles alors que les adultes recherchent les eaux plus froides. Ceci est très net quand l'été est chaud et que le gradient thermique est fort dans l'épilimnion. Dans ce cas, les relations de prédation inter- et intraspécifiques sont réduites. Ce n'est plus le cas à l'automne et en hiver mais, à cette époque, les jeunes constituent des proies plus mobiles qui ont une meilleure chance de survie.
III. CONCLUSION
Parler de peuplement piscicole est une solution de facilité pour «compartimenter» I'écosystème mais ne recouvre pas une entité fonctionnelle réelle en ce qui concerne les peuplements des lacs tempérés du moins. Ces peuplements ont une richesse spécifique généralement très faible ; de ce fait, il existe sans doute moins de relations entre les composantes du peuplement piscicole qu'entre le reste de l'écosystème. La structure spécifique actuelle est souvent le fruit d'acclimatations plus ou moins réussies. La dynamique de chaque population est beaucoup plus influencée par les paramètres du milieu extérieurs au compartiment pisciaire que par des relations intraspécifiques des composantes du peuplement.
Les commentaires sont fermés.