
Adaptation aux eaux douces
01/03/2011
Les animaux d'eau douce (dulçaquicoles) ont soit une origine marine, soit une origine terrestre. Dans le premier cas, ils auront à résoudre un problème de pression osmotique. Dans le second cas, ils auront à résoudre un problème de respiration.
Physiologie de l'adaptation aux eaux douces
par André Guyard
Il ne semble exister aucun groupe d'organismes pluricellulaires d'origine dulçaquicole sauf peut-être quelques lignées de Turbellariés. Tous les groupes actuels d'organismes dulçaquicoles semblent avoir
- une origine marine [Éponges, Cnidaires, Aschelminthes (Némathelminthes + Rotifères), Annélides, Mollusques, Crustacés et Vertébrés inférieurs] ;
- ou une origine terrestre : Plantes à fleurs, Insectes, Arachnides et Vertébrés Amniotes. En conséquence, selon l'origine du groupe, les problèmes physiologiques de l'invasion des eaux douces sont totalement différents.
I. Invasions d'origine marine
Les colonisations marines sont distribuées irrégulièrement et de façon dispersée dans le règne animal. Certains groupes comme les Échinodermes, les Céphalopodes et probablement les Brachiopodes ont été totalement incapables de s'adapter aux eaux douces.

Namanereis sp. © Calphoto Berkeley/Gustav Paulay
Parmi les Polychètes, seuls quelques genres de la famille des Nereidae apparentés à Namanereis (=Lycastis) sont susceptibles de s'adapter à l'eau douce.
Il est tentant de supposer que certains groupes sont prédisposés à la vie en eau douce. Cette préadaptation porterait sur les capacités suivantes :
1. Préadaptations à la vie en eau douce : contrôle du paramètre pression osmotique
- 1. 1. perméabilité à l'eau et aux électrolytes inférieure à la moyenne ;
- 1.2. capacité d'accomplir le travail cellulaire sous une pression osmotique inférieure à celle de l'eau de mer ;
- 1.3. capacité de maintenir une proportion ionique correcte dans les cellules soumises à des concentrations variables du fluide circulant ;
- 1.4. capacité d'excrétion de l'eau pour maintenir une pression osmotique constante ;
- 1.5. capacité de développer un mécanisme d'absorption des sels contre un gradient de diffusion.
Il apparaîtra dans la discussion que la première de ces caractéristiques est la plus importante. Mais au cours de l'évolution, une cinquième faculté, peut-être une spécialisation de la troisième (excrétion de l'eau), développée seulement plus tard dans le processus de l'adaptation, est la possibilité de sécréter une urine hypotonique par rapport au sang. (cette faculté devient une importante condition quand la perméabilité de la paroi du corps à l'eau reste élevée).
Outre la pression osmotique, d'autres facteurs d'adaptation peuvent être évoqués. Il est évident que la variabilité de la température des eaux douces peut constituer une barrière thermique importante aux migrations eau de mer-eau douce. Il y a aussi une bonne raison de penser que l'ontogenèse avec larves pélagiques, si répandue dans le milieu marin, est inadéquate en eau douce.
2. Le problème osmotique
Tous les Invertébrés marins ont une pression osmotique voisine de celle de l'eau de mer. On dit qu'ils sont strictement poïkilosmotiques. Un passage brutal dans de l'eau douce entraîne une dilution du milieu intérieur incompatible avec le maintien de l'intégrité des mécanismes biochimiques. Pour passer en eau douce, un degré considérable d'homéosmotisme doit être développé.
2.1. L'ionorégulation
Dans les organismes marins, le taux de potassium intracellulaire est plus grand que le taux de potassium extracellulaire. C'est le contraire pour les ions SO42- et Mg2+. Le rapport intracellulaire K+/Na+ le plus haut est requis pour les cellules contractiles et conductrices. La régulation ionique existe dans tout le règne animal depuis les Cnidaires. Elle se fait à travers le tégument, au niveau de l'appareil excréteur, respiratoire ou digestif. Elle constitue la base de l'osmorégulation.
2.2. Adaptations euryhalines
Vivnat dans la zone intertidale et subissant des émersions quotidiennes, certains animaux marins sont plus euryhalins que d'autres. Arenicola, Macoma baltica et Mytilus edulis sont poïkilosmotiques et euryhalins. Des animaux marins supportent des variations de 25 à 40 ‰ de la salinité. Les organismes des estuaires sont typiquement euryhalins : Marinogammarus obtusatus, Nereis diversicolor, Mya arenaria. La tendance à l'euryhalinité va de pair avec le développement d'un tégument perméable et des surfaces respiratoires. La perméabilité à l'O2 va de pair avec la perméabilité à l'eau. Il est probable que deux types différents d'adaptations se sont produites.

Mya arenaria ©galeriabaltyk

Macoma baltica (cliché DR)
Marinogammarus obtusatus

Nereis diversicolor © Ricardo R. Fernandez
2.3. Solutions choisies par les animaux dulçaquicoles pour résoudre le problème osmotique
- 2.3.1. L'option Lamellibranche : dilution du milieu interne
Chez les Lamellibranches, où une grande surface de cténidies est exposée au milieu externe, l'adaptation consiste en une dilution importante du milieu interne (20-46 mM/l) et une dilution plus modérée du milieu intracellulaire. Chez Anodonta, genre ayant la plus faible concentration sanguine connue dans le règne animal, la concentration intracellulaire est égale à 1,5 fois la concentration du milieu intérieur. Dans ce cas les cellules travaillent dans des concentrations d'ions et d'autres substances actives osmotiquement inhabituellement basses. L'eau qui rentre dans les tissus est excrétée sous forme d'une urine abondante et hypotonique par rapport au sang. La production journalière équivaut au quart du poids de l'animal à 15-18 °C.

Anodonta cygnea © F. Brabec
2.3.2. L'option crabe d'eau douce : forte concentration sanguine et urine isotonique et peu abondante
Chez les crabes d'eau douce où la surface branchiale est réduite et le corps recouvert d'un exosquelette chitineux et chez des formes d'eau saumâtre chez lesquelles une relativement basse perméabilité à l'eau et aux sels couplée avec une forte capacité d'absorption active d'ions à partir de l'environnement, l'adaptation consiste en une forte concentration sanguine couplée avec l'émission d'une urine quasi isotonique et peu abondante.

Crabe d'eau douce : Eriocheir sinensis © Mattdufort
Potamon niloticus peut survivre 3 semaines dans de l'eau de mer diluée à 50 %, 1 à 3 semaines dans de l'eau de mer diluée à 25 % et pas plus de 4 jours dans de l'eau de mer pure. La concentration sanguine de Potamon niloticus atteint 280 mM dans de l'eau de mer à 50 % et croît rapidement. L'animal moribond dans l'eau de mer est approximativement isotonique avec le milieu. La mort est due à la déshydratation de certains tissus, en particulier musculaires où la teneur en eau chute à 25 %. Dans ces conditions, on note un faible accroissement du sang en acides aminés et une montée du sodium mais ces accroissements sont insuffisants pour assurer l'équilibre osmotique du tissu musculaire avec le sang. Le Crabe enragé Carcinus maenas transféré dans de l'eau de mer plus concentrée, libère une plus grande quantité d'acides aminés dans ses muscles et ainsi contrôle mieux son contenu aqueux. La production d'urine est faible (moins de 1 % du poids du corps par jour), alors que la production journalière d'urine chez Anodonta équivaut au 1/4 du poids à température ordinaire (15-18°C).
Mis à part les crabes fluviatiles, les Gammaridae, les Astacidae et les moules d'eau douce, nous ne connaissons pas la répartition systématique de ces deux modes extrêmes d'adaptation. Le fait est intéressant du point de vue de l'écologie évolutive. On pourrait suspecter que le type d'adaptation présenté par Anodonta, dans lequel milieu externe, milieu interne et tissus montrent une pression osmotique décroissant parallèlement, serait une adaptation passive à la vie en eau douce, alors que le type d'adaptation montré par Potamon serait actif et traduirait une adaptation à des eaux saumâtres de salinité variable. Les stades initiaux seraient plus difficiles pour un animal avec une énorme surface épithéliale que pour un animal protégé par un exosquelette même si ce dernier est loin d'être imperméable. Ceci expliquerait le plus grand nombre d'invasions des eaux douces réussies par les Crustacés que par les Mollusques qui constituent pourtant un groupe plus nombreux en espèces marines.
Quel que soit le cours de l'évolution, le milieu interne des animaux dulçaquicoles n'est jamais aussi dilué que le milieu extérieur et la paroi du corps jamais complètement imperméable à l'eau, de sorte qu'un gradient osmotique existe toujours avec un flux entrant d'eau. Le problème physiologique qui se pose est la capture des sels dans un milieu hypotonique et leur rétention. La régulation du flux entrant d'eau est calcium-dépendante. Chez les animaux qui perdent du sel dans l'eau d'excrétion et chez les animaux en cours de croissance, un mécanisme de capture des ions contre le gradient de concentration est nécessaire (pompes actives). Dans certains cas, le chlorure de sodium alimentaire suffit à l'animal, mais les organes de capture d'ions dans le milieu sont bien développés notamment en rapport avec les surfaces respiratoires. Il est probable qu'un grand nombre d'animaux, notamment les Crustacés, qui ont résolu leur problème osmotique sont incapables de s'avancer en milieu continental loin de la côte à cause de leurs besoins de quelques mg de chlorure/l dans le milieu. De même, un accroissement de la température accroît le taux de consommation d'eau et d'excrétion. Il est probable que ces mécanismes fonctionnent mieux à haute température et que la pénétration d'une espèce en eau douce est limitée par les températures froides hivernales.
Parmi les vertébrés inférieurs, les Myxinoïdes sont probablement passés par un stade en eau douce chez leurs ancêtres, les espèces retournant à la mer ayant conservé une concentration d'électrolytes sanguins inférieure à celle de l'eau de mer. Chez les Élasmobranches, le déficit en concentration osmotique est compensé par de l'urée et de la triméthylamineoxyde. Les autres Poissons marins ont un sang hypotonique. Ils conservent l'eau de boisson et excrètent du chlorure à travers leurs branchies. Ce mécanisme maintient la concentration sanguine plus basse que celle de l'eau de mer mais deux fois plus grande que celle des poissons d'eau douce.
Certains poissons migrant de l'eau de mer vers l'eau douce ou vice-versa sont capables de changer leur concentration sanguine. Cette faculté est sous la dépendance d'un système de contrôle endocrine notamment par la sécrétion pituitaire de prolactine. Les Amniotes, Reptiles et Oiseaux, peuvent être secondairement associés au milieu aquatique marin ou d'eau douce. En eau de mer, l'excès de sel absorbé avec la nourriture est excrété par la glande orbitaire.
II. Invasions d'origine terrestre
Les problèmes rencontrés par une espèce terrestre s'adaptant à la vie dans les eaux douces sont complètement différents de ceux rencontrés par une espèce marine. Il ne s'agit plus de problèmes de pression osmotique mais de respiration. De nombreux animaux aquatiques continuent à respirer l'oxygène gazeux sous l'eau en remontant régulièrement à la surface pour respirer, en restant en contact avec l'atmosphère grâce à un siphon ou en emportant une provision d'air avec eux. Dans cette dernière option, les cas les plus remarquables sont fournis par les plastrons respiratoires des insectes aquatiques. Mise à part cette adaptation, les organismes aquatiques développent un appareil respiratoire branchial, permettant l'utilisation directe de l'oxygène dissous dans l'eau.
1. Gaz respiratoires : comparaison entre animaux terrestres et animaux aquatiques
On sait depuis Lavoisier que les animaux terrestres prélèvent de l'oxygène dans l'air et y rejettent du CO2 et que la chaleur animale est produite par un phénomène chimique analogue à la combustion. C'est à un physiologiste italien, Spallanzani que l'on doit la découverte de la respiration chez les animaux aquatiques.
1.1. L'O2
Les métabolismes producteurs d'énergie utilisent l'O2 comme comburant et comme carburant des substances organiques : glucides, lipides, protides. Le résultat de ces métabolismes est la production de CO2 et d'eau.
La variation des quantités d'O2 et de CO2 dans le milieu respiré par les animaux s'accompagne d'une variation de la concentration de ces gaz et par conséquent de leurs pressions partielles encore appelées tensions. Il existe en effet, pour chaque corps chimique, une relation entre sa concentration dans l'eau ou dans l'air et sa pression partielle. Lorsque l'eau et l'air sont en équilibre, les pressions partielles dans chacun de ces milieux sont identiques mais les concentrations d'oxygène sont très différentes, l'air contenant beaucoup plus de molécules d'oxygène que l'eau. En quelque sorte, on peut dire que l'oxygène est plus « soluble » dans l'air que dans l'eau. La diffusion d'un gaz à travers un gaz est plus facile qu'à travers un liquide et la pression partielle d'oxygène par litre d'air est beaucoup plus importante que par litre d'eau de sorte que la respiration aérienne est plus efficace que la respiration de l'oxygène dissous. On appelle capacitance (solubilité) la relation qui existe entre la variation de la concentration d'un gaz dans un milieu et la variation de sa pression partielle. La capacitance se définit comme le rapport de ces deux quantités. Elle mesure la capacité d'un milieu à contenir un gaz en fonction de la pression partielle de ce gaz. Par comparaison avec l'air, les animaux aquatiques peuplent un milieu relativement pauvre en oxygène (O2 est environ 30 fois moins soluble dans l'eau que dans l'air) et où les pressions partielles, PO2 et de P CO2 sont extrêmement diverses et variables. D'après Dejours (1981), dans des conditions particulières, la PO2 peut varier de 0 à 300 Torr (mm Hg) et la PCO2 de 0 à 10-15 Torr. Rappelons que dans l'air, au niveau de la mer, PO2 est sensiblement égale à 155 Torr et PCO2 à 0,25 Torr.
Selon qu'une quantité d'oxygène donnée est prelevée dans l'eau ou bien dans l'air, les variations de pressions partielles dans ces deux milieux sont très différentes. Ainsi, quand un animal aquatique abaisse de 50 Torr la pression d'oxygène dans l'eau qu'il exhale, il prélève 1/10 de millimole d'oxygène dans chaque litre d'eau respiré. L'animal aquatique doit ainsi inhaler 10 l d'eau pour en soustraire 1 millimole d'O2 qui diffuse dans le sang [ventilation spécifique (équivalent respiratoire) = 10 l/millimole d'O2]. Un saumon de quelques Kg qui consomme 1 mole d'O2 par jour doit respirer une dizaine de m3 d'eau pendant le même temps. En revanche, un animal aérien qui respire l'air qui, à pression partielle égale, est beaucoup plus riche en O2 que l'eau, est capable de prélever par litre d'air 1,67 millimole d'oxygène lequel diffuse dans le sang par les membranes alvéolaires des poumons. Sa ventilation spécifique vaut 1/1,67 = O,6 l/millimole d'O2 alors que chez le poisson, elle est de 10 l/millimole d'O2.
1.2. Le CO2
L'autre gaz respiratoire est le CO2. Contrairement à ce qui se passe dans l'air, la solubilité (capacitance) du CO2 dans l'eau est beaucoup plus élevée que celle de l'O2. L'addition dans l'eau de 0,1 mmole de CO2 s'accompagne d'une variation de pression partielle beaucoup plus faible que celle qui correspond au prélèvement de la même quantité d'O2. C'est pourquoi le CO2 est facilement rejeté dans l'eau et sa pression partielle n'est jamais très élevée dans le sang de l'animal aquatique. Elle n'atteint que 1 ou 2 Torr dans un animal aquatique vivant dans une eau bien aérée. Du fait de la respiration, la diminution de la PO2 dans l'eau ou dans l'air inhalés et exhalés, est du même ordre de grandeur chez le poisson et chez le canard tandis que l'élévation de PCO2 est de 1 à 2 Torr chez l'animal aquatique mais atteint plusieurs dizaines de Torr chez l'animal aérien. En bref, l'animal aquatique qui respire un milieu plus pauvre en O2 ventile beaucoup et la PCO2 dans l'eau exhalée est basse tandis que l'animal aérien qui se trouve dans un milieu riche en O2 ventile peu et la PCO2 de l'air exhalé est élevée.
Comme la viscosité et la densité de l'eau sont respectivement 60 et 800 fois plus élevées que celles de l'air, le travail respiratoire de l'animal aquatique est considérable. 10 à 20 % de la dépense totale d'énergie d'un poisson sont utilisés pour la respiration externe.
En fait, quel que soit le mode de respiration, l'oxygène respiré passe toujours par une phase en solution dans l'eau. Si l'espèce aquatique est normalement munie d'un appareil respiratoire aquatique, l'espèce terrestre qui est recouverte d'un tégument imperméable pour prévenir l'évaporation, possède un épithélium respiratoire enfermé dans des cavités poumons ou trachées, espaces facilement saturées en vapeur d'eau.
2. Respiration et alcalinité du sang
2.1. Effets de l'oxygène dissous
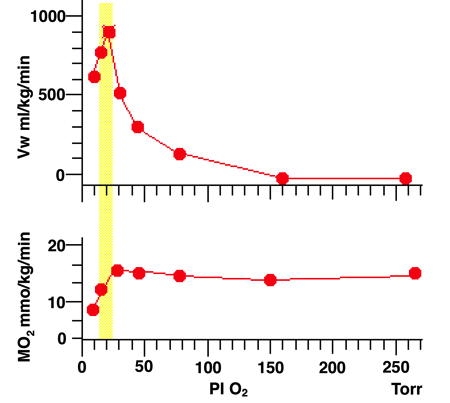
La figure ci-dessus montre l'évolution du débit ventilatoire, Vw et de la consommation d'oxygène, MO2, chez l'Écrevisse Astacus leptodactylus exposée à différentes valeurs de la pression partielle d'oxygène dans l'eau inspirée. PI O2. De 240 à 50 Torr, MO2 est maintenue constante au prix d'une multiplication par 20 du débit ventilatoire. La zone pointillée indique la valeur de PO2 au-dessous de laquelle MO2 ne peut plus être maintenue (température 13°C, pression partielle de CO2 : 0,8 Torr) (D'après P. Dejours, 1983).
En raison de sa faible concentration dans l'eau, l'approvisionnement en O2 tient un rôle privilégié dans les adaptations ventilatoires des animaux aquatiques. Ceux-ci, pour maintenir leur métabolisme aérobie, doivent ajuster d'une facon étroite leur ventilation à la quantité d'oxygène disponible. La figure 1 illustre cette relation générale chez l'Écrevisse, Astacus leptodactylus . Dans une très large gamme de PO2, le débit ventilatoire est d'autant plus important que le niveau d'oxygénation est faible. Cette dépendance vis-à-vis de l'oxygène a d'importantes conséquences sur la composition du milieu intérieur. En effet, les échanges respiratoires déterminent non seulement l'apport de l'oxygène aux cellules, mais également l'élimination du CO2 produit par le métabolisme cellulaire. Une diminution de la vitesse de ces échanges, comme on l'observe en hyperoxie, a pour conséquence une accumulation de CO2 et une acidification du milieu extracellulaire. Or chez l'animal aquatique, contrairement à ce qu'on observe chez l'animal aérien, l'augmentation de la PCO2 en hyperoxie ne s'accompagne pas d'une stimulation respiratoire. La ventilation branchiale semble ici principalement règlée en vue d'assurer la fourniture d'oxygène aux tissus et elle est peu ou pas influencée par le CO2 .
2.2. Effets du CO2
L'effet du CO2 est encore un problème fort controversé car l'effet du CO2 dépend de l'état physiologique des animaux. Chez les Crustacés, le niveau métabolique peut varier d'un facteur 2 entre le jour et la nuit, période où l'animal est actif et d'un facteur 12 entre les périodes de repos hivernal et celles de pleine activité estivale. D'après Massabuau (1981), la ventilation est bien stimulée par le CO2 en normoxie, mais la réaction est liée à la fois au niveau métabolique et au niveau d'oxygénation. La figure 2 illustre cette dépendance vis-à-vis du métabolisme en utilisant les variations spontanées d'activités que l'on observe au niveau du nycthémère. Des écrevisses normoxiques (dans un milieu dont la PO2 est identique à celle observée dans l'air, faiblement réactives au CO2 au cours de la journée ont été exposées à des eaux hypercapniques (PCO2 supérieure à celle observée dans l'air), soit le matin (période où l'animal est au repos), soit le soir (période ou l'animal est actif).
Alors que le jour, la ventilation est peu ou pas affectée, la nuit l'inhalation d'eau hypercapnique stimule l'activité ventilatoire. De plus, lorsque ces expositions hypercapniques sont réalisées alors que l'animal baigne dans une eau hyperoxique, les réponses au CO2 sont faibles ou nulles comme Dejours (1981) l'a montré chez la Truite .
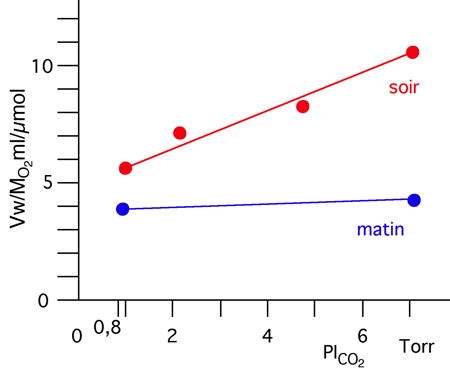
La figure ci-dessus montre l'évolution du débit ventilatoire nécessaire à l'extraction d'une quantité unitaire d'oxygène, Ww/MO2, en fonction de la pression partielle de CO2 dans l'eau inspirée PICO2 et du rythme circadien d'activité (chez des écrevisses A. leptodactylus choisies pour leur faible réponse le matin en normoxie à 13°C). Le soir, alors que les animaux sont actifs, le CO2 stimule clairement la ventilation. (D'après P. Dejours, 1983).
Il existe donc une régulation ventilatoire des échanges de CO2 chez les animaux aquatiques. La branchie est non seulement un échangeur de gaz mais également un échangeur d'ions.
3. Autres différences entre animaux aquatiques et animaux aériens
Chez l'animal aquatique, en vertu de la poussée d'Archimède, la pression mécanique de l'eau est équivalente à la pression artérielle, car la densité du sang est à peu près équivalente à la densité de l'eau. De sorte qu'il n'est pas nécessaire que la paroi des vaisseaux sanguins soit épaisse.
De même, le poids du squelette des animaux aquatiques représente une faible part du poids total et cette part est indépendante de la taille de l'animal.
En ce qui concerne la chaleur interne, elle est rapidement dissipée chez les animaux aquatiques puisque la capacité et la conductivité caloriques de l'eau sont élevées. Les animaux aquatiques ont à peu près la même température que l'eau dans laquelle ils vivent. Leur température leur est imposée par celle de l'eau. Ils sont poïkilothermes. Les animaux aquatiques sténothermes qui, par définition ont une température corporelle qui ne varie guère, ne maintiennent celle-ci qu'en vivant dans des eaux dont la température leur est propice.
Si l'on excepte l'adaptation du contrôle des mouvements d'ions, d'eau et d'oxygène, l'environnement aquatique d'eau douce impose certainement d'autres conditions aux organismes colonisateurs. La température variable des eaux douces dans les régions tempérées constituerait une barrière non négligeable à leur invasion par les organismes marins, mais le grand nombre de formes dans les eaux hypogées qui descendent de formes ancestrales marines suggère que la stabilité thermique des cavités souterraines a grandement facilité le processus. L'invasion des eaux hypogées a dû se faire selon deux voies :
- une voie passive dans des grottes sous-marines progressivement isolées et alimentées en eau douce ;
- une voie active par les eaux phréatiques.
On peut supposer que les groupes dont le développement exige la présence de petits stades larvaires planctoniques ont des difficultés à envahir les eaux douces. En effet, il est vraisemblable que la survie de stades aussi précoces dans les eaux courantes est fortement compromise à cause de leur passivité vis-à-vis des courants et de leur densité vis-à-vis des eaux douces. En général, il semble que les organismes de grande taille ont plus de difficultés à s'adapter aux eaux douces que les petits organismes. De plus, souvent, les races adaptées genétiquement à différentes salinités sont morphologiquement indistinguables. C'est le cas notamment de la famille des Gammaridae. Il est probable que l'adaptation des Gammaridae à la vie en eau douce est en cours à l'ère actuelle.

Gammarus pulex © Wontolla
Les commentaires sont fermés.